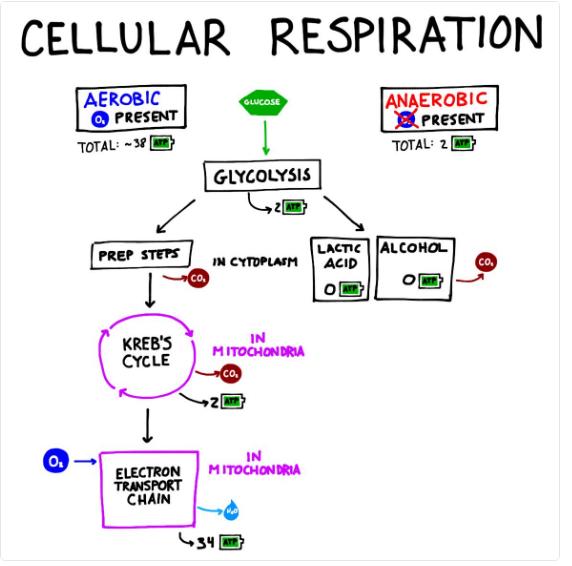
Oxidative Phosphorylation: Steps, Location, ATP Yield & Mechanism
Every living cell requires a constant supply of energy to perform essential functions such as growth, repair, and active transport. This energy is primarily stored and transferred in the form of ATP (adenosine triphosphate), often referred to as the “energy currency” of the cell. Without ATP, vital cellular processes would come to a halt, making energy production a central aspect of life.
Cells generate ATP through a series of metabolic pathways collectively known as cellular respiration. This multi-step process includes glycolysis, the Krebs cycle, and the final stage—oxidative phosphorylation. Among these stages, oxidative phosphorylation is the most efficient and produces the majority of ATP required by the cell.
In this process, high-energy electrons derived from NADH and FADH₂ are transferred through the electron transport chain, ultimately driving ATP synthesis. Understanding oxidative phosphorylation is crucial for students and researchers alike, as it explains how cells harness energy from nutrients to sustain life.
What is oxidative phosphorylation?
Oxidative phosphorylation is the process by which cells generate ATP using electrons transferred through the electron transport chain to create a proton gradient that drives ATP synthesis.
Quick Facts About Oxidative Phosphorylation
| Feature | Details |
| Location | Inner mitochondrial membrane |
| Inputs | NADH, FADH₂, O₂ |
| Outputs | ATP, H₂O |
| Key Components | Electron Transport Chain + Chemiosmosis |
| Oxygen Role | Final electron acceptor |
| ATP Yield | ~26–28 ATP per glucose |
Oxidative phosphorylation is the final stage of cellular respiration and is responsible for producing the largest amount of ATP. It combines two key processes—the Electron Transport Chain and chemiosmosis—to efficiently convert energy stored in electrons into usable cellular energy in the form of ATP.
What is Oxidative Phosphorylation?
Oxidative phosphorylation is a biochemical process in which cells produce ATP by transferring electrons through a series of protein complexes embedded in the inner mitochondrial membrane. In simple terms, it is the stage of cellular respiration where most of the cell’s usable energy is generated.
As the final step of cellular respiration, oxidative phosphorylation follows glycolysis and the Krebs cycle. During these earlier stages, high-energy electron carriers such as NADH and FADH₂ are produced. These molecules then donate their electrons to the Electron Transport Chain, which plays a central role in driving ATP production.
The energy released as electrons move through the electron transport chain is used to pump protons (H⁺ ions) across the inner mitochondrial membrane, creating a proton gradient. This gradient stores potential energy, which is later utilized by ATP synthase to convert ADP into ATP. Through this mechanism, oxidative phosphorylation becomes the primary source of ATP in aerobic organisms.
Overall, oxidative phosphorylation is essential for efficient energy production, enabling cells to meet their metabolic demands and sustain vital biological functions.
Where Does Oxidative Phosphorylation Occur?
Oxidative phosphorylation takes place inside the mitochondria, specifically along the inner mitochondrial membrane. This location is highly specialized and perfectly suited for efficient ATP production.
The inner mitochondrial membrane is folded into structures called cristae, which significantly increase the surface area available for biochemical reactions. These folds house the protein complexes of the Electron Transport Chain and the enzyme ATP Synthase, both of which are essential for oxidative phosphorylation.
This membrane also creates two distinct compartments:
- The matrix (inside the mitochondria)
- The intermembrane space (between outer and inner membranes)
As electrons pass through the electron transport chain, protons (H⁺ ions) are pumped from the matrix into the intermembrane space. This results in the formation of a proton gradient across the inner membrane. The stored energy in this gradient is then used by ATP synthase to generate ATP.
Because of this highly organized structure and compartmentalization, the mitochondria are often referred to as the “powerhouse of the cell,” with oxidative phosphorylation being the key process responsible for large-scale ATP production.
Steps of Oxidative Phosphorylation
Oxidative phosphorylation is a multi-step process that efficiently converts energy stored in electron carriers into ATP. It consists of three major stages: the Electron Transport Chain, proton gradient formation, and ATP synthesis through chemiosmosis.
1. Electron Transport Chain (ETC)
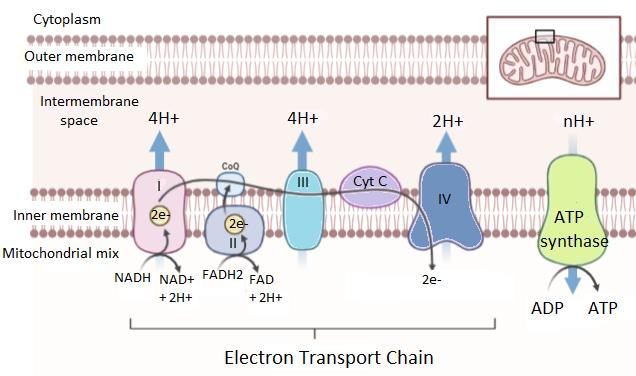
The electron transport chain is the first and most crucial step of oxidative phosphorylation. It is a series of protein complexes (Complex I, II, III, and IV) located in the inner mitochondrial membrane.
- NADH and FADH₂, produced during glycolysis and the Krebs Cycle, donate high-energy electrons to the ETC.
- NADH transfers electrons to Complex I, while FADH₂ donates electrons to Complex II.
- These electrons move through a chain of carriers, including coenzyme Q and cytochrome c.
As electrons pass through these complexes, energy is released step by step. This energy is not used directly to make ATP but instead powers the pumping of protons across the membrane.
Finally, oxygen acts as the terminal electron acceptor, combining with electrons and protons to form water. This step is essential—without oxygen, the entire process would stop
2. Proton Gradient Formation
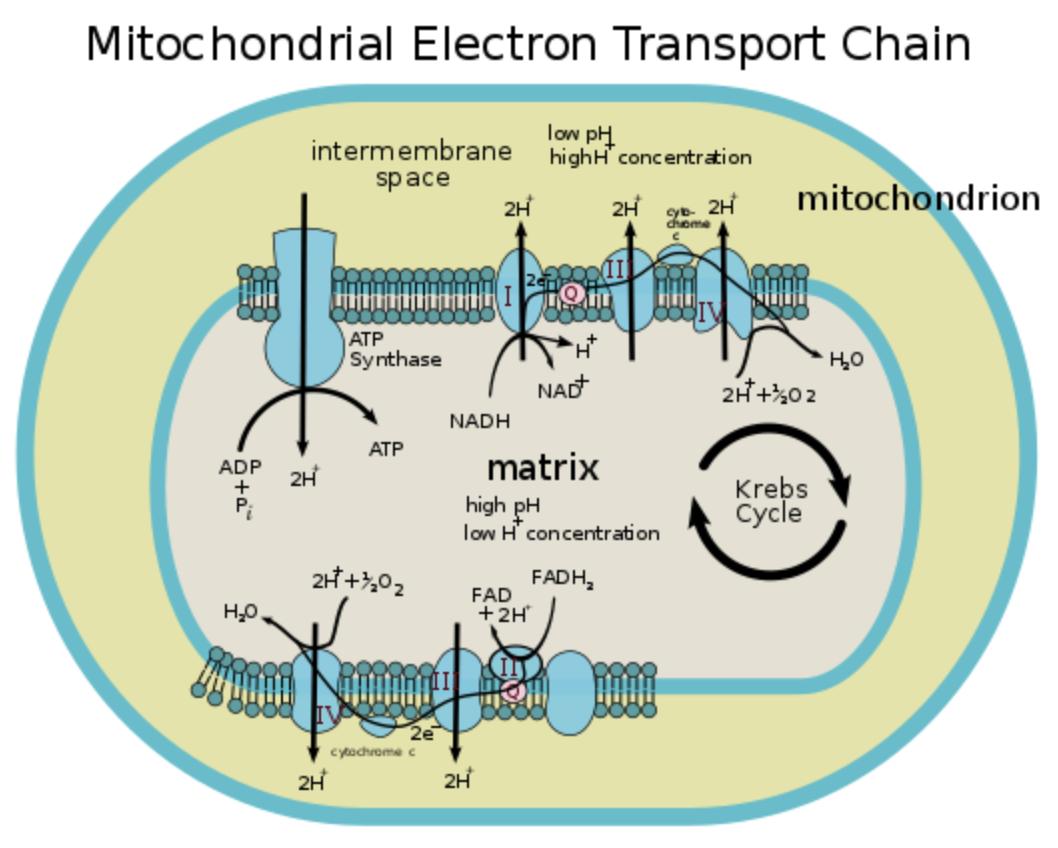
As electrons move through the electron transport chain, the released energy is used to pump protons (H⁺ ions) from the mitochondrial matrix into the intermembrane space.
- Complexes I, III, and IV actively pump protons across the membrane.
- This creates a high concentration of protons in the intermembrane space and a low concentration in the matrix.
This difference in proton concentration generates an electrochemical gradient, also known as the proton motive force. It stores potential energy, similar to water held behind a dam.
This gradient is the key driving force behind ATP production in the next step.
3. ATP Synthesis (Chemiosmosis)
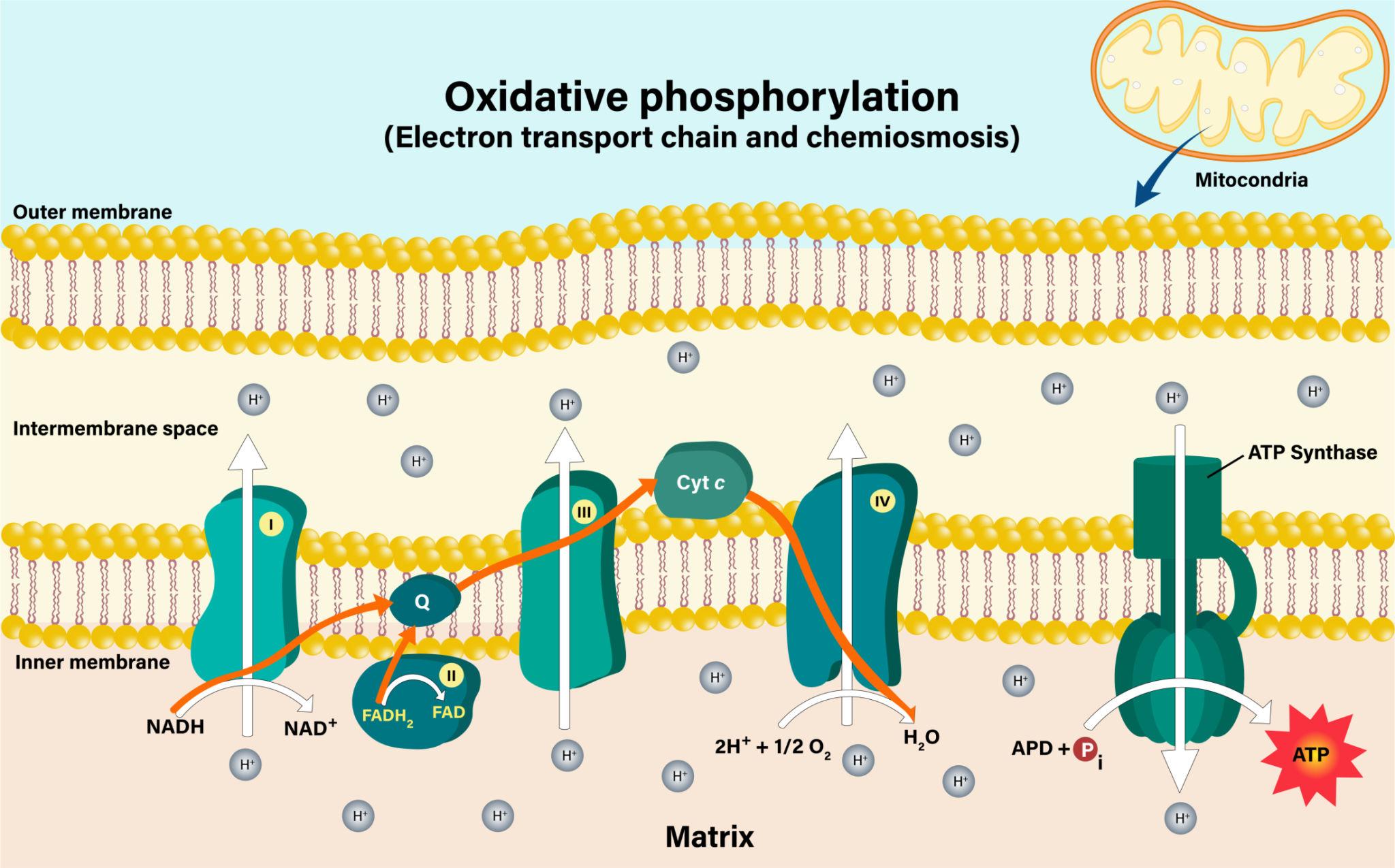
The final step of oxidative phosphorylation is ATP synthesis through chemiosmosis.
- Protons flow back into the mitochondrial matrix through the enzyme ATP Synthase.
- This flow of protons provides the energy needed to drive the conversion of ADP (adenosine diphosphate) into ATP.
ATP synthase functions like a molecular turbine. As protons pass through it, the enzyme rotates and catalyzes the formation of ATP from ADP and inorganic phosphate (Pi).
This process produces the majority of ATP in aerobic respiration, making oxidative phosphorylation the most efficient energy-generating stage in the cell.
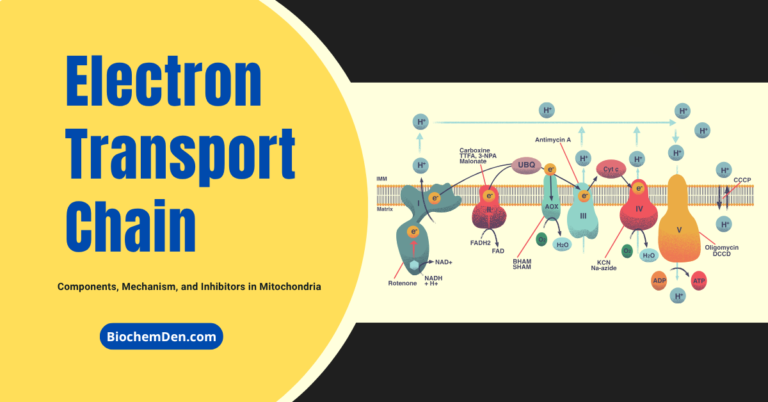
Electron Transport Chain
The Electron Transport Chain (ETC) is the core component of oxidative phosphorylation, where high-energy electrons from NADH and FADH₂ are transferred through a series of protein complexes to ultimately generate ATP. This process occurs in the inner mitochondrial membrane and is responsible for establishing the proton gradient that drives chemiosmosis.
Structure and Components of the Electron Transport Chain
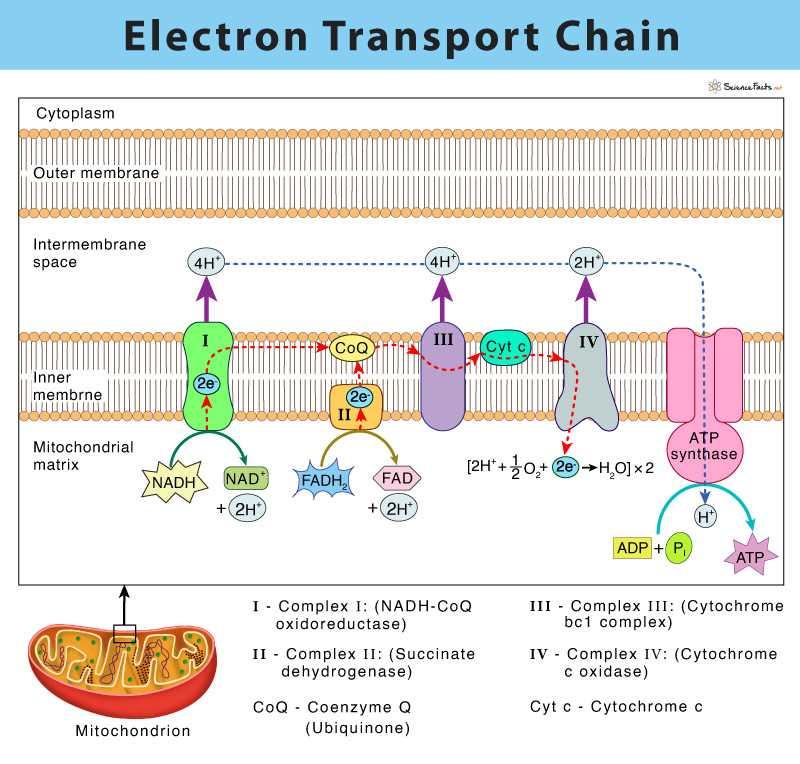
The electron transport chain consists of four major protein complexes (Complex I–IV) and two mobile electron carriers:
- Coenzyme Q (Ubiquinone)
- Cytochrome c
These components work together to ensure a smooth and efficient flow of electrons.
Complex I (NADH Dehydrogenase)
- Receives electrons from NADH
- Transfers electrons to coenzyme Q
- Pumps protons (H⁺) from the matrix to the intermembrane space
This complex plays a major role in establishing the proton gradient.
Complex II (Succinate Dehydrogenase)
- Accepts electrons from FADH₂
- Transfers them to coenzyme Q
- Does not pump protons, unlike other complexes
Because it doesn’t contribute to proton pumping, FADH₂ generates less ATP compared to NADH.
Role of Coenzyme Q (Ubiquinone)
- A lipid-soluble carrier within the membrane
- Transfers electrons from Complex I and II to Complex III
It acts as a central hub for electron transfer in the ETC.
Complex III (Cytochrome bc₁ Complex)
- Receives electrons from coenzyme Q
- Transfers them to cytochrome c
- Pumps protons across the membrane
This step further strengthens the proton gradient.
Role of Cytochrome c
- A small protein located in the intermembrane space
- Transfers electrons from Complex III to Complex IV
It ensures efficient electron movement between these complexes.
Complex IV (Cytochrome c Oxidase)
- Accepts electrons from cytochrome c
- Transfers them to oxygen (O₂), the final electron acceptor
- Pumps protons across the membrane
This is the final step in the electron transport chain.
Oxygen as the Final Electron Acceptor
Oxygen plays a critical role in oxidative phosphorylation. At Complex IV:
- Oxygen accepts electrons and combines with protons (H⁺)
- This reaction forms water (H₂O)
Without oxygen, electrons cannot be removed from the system, causing the entire electron transport chain to stop. This is why oxidative phosphorylation only occurs under aerobic conditions.
Formation of Water
At the end of the electron transport chain:
- Electrons + Oxygen + Protons → Water (H₂O)
This reaction not only removes electrons from the chain but also maintains the continuous flow of electrons, ensuring ATP production.
Role of Oxygen in Oxidative Phosphorylation
Oxygen plays a decisive role in oxidative phosphorylation by ensuring the continuous flow of electrons through the electron transport chain. Without oxygen, this entire energy-generating process would come to a halt.
Why is Oxygen Essential?
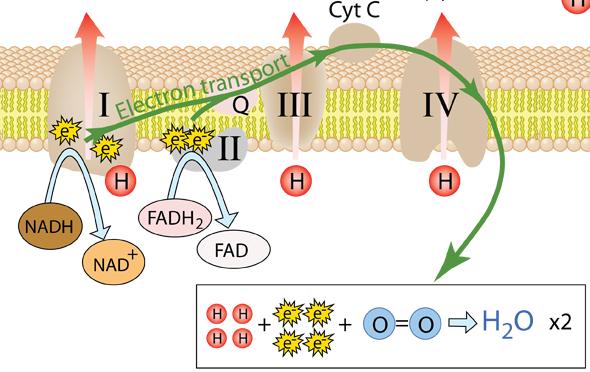
Oxygen acts as the final electron acceptor at the end of the electron transport chain (Complex IV).
- Electrons travel through complexes I–IV and reach oxygen
- Oxygen combines with electrons and protons (H⁺) to form water (H₂O)
- This step prevents the accumulation of electrons in the chain
By accepting electrons, oxygen allows the ETC to continue functioning smoothly. This continuous electron flow is essential for maintaining the proton gradient that drives ATP synthesis via chemiosmosis.
What Happens Without Oxygen?
In the absence of oxygen (anaerobic conditions):
- The electron transport chain becomes blocked
- NADH and FADH₂ cannot donate electrons efficiently
- Proton pumping stops, collapsing the proton gradient
- ATP production through oxidative phosphorylation ceases
As a result, cells are forced to rely on less efficient pathways to generate energy.
Link to Anaerobic Respiration
When oxygen is unavailable, cells shift from oxidative phosphorylation to anaerobic respiration.
- Energy is produced mainly through glycolysis
- NAD⁺ is regenerated via fermentation (e.g., lactic acid fermentation)
- Only a small amount of ATP (2 ATP per glucose) is generated
Although anaerobic respiration allows cells to survive temporarily without oxygen, it is far less efficient compared to oxidative phosphorylation.
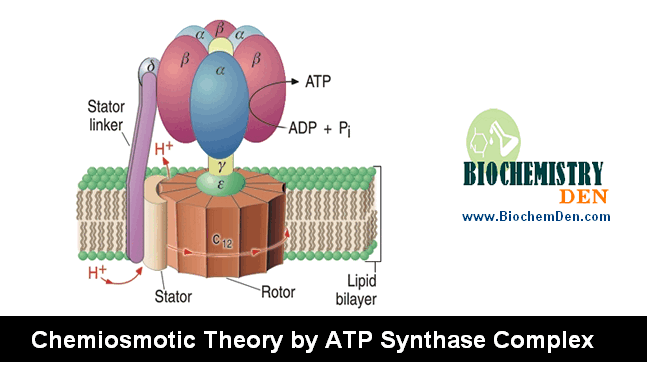
Chemiosmosis and ATP Synthase Mechanism
Chemiosmosis is the final stage of Oxidative Phosphorylation, where the energy stored in a proton gradient is converted into ATP. This process links the Electron Transport Chain to ATP production and is essential for efficient cellular energy generation.
Proton Motive Force (PMF)
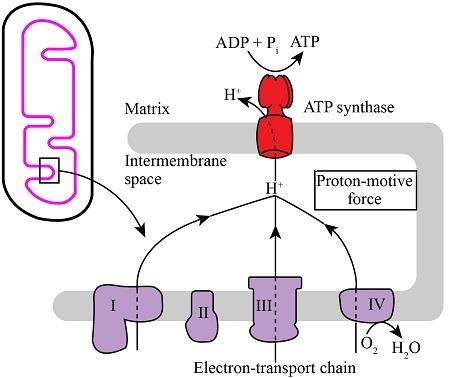
During electron transport, protons (H⁺ ions) are pumped from the mitochondrial matrix into the intermembrane space by complexes I, III, and IV. This creates an electrochemical gradient known as the proton motive force (PMF).
The proton motive force has two components:
- Chemical gradient (ΔpH): Difference in proton concentration
- Electrical gradient (Δψ): Difference in charge across the membrane
This gradient stores potential energy, similar to water held behind a dam, and is the driving force behind ATP synthesis.
ATP Synthase Rotation Mechanism
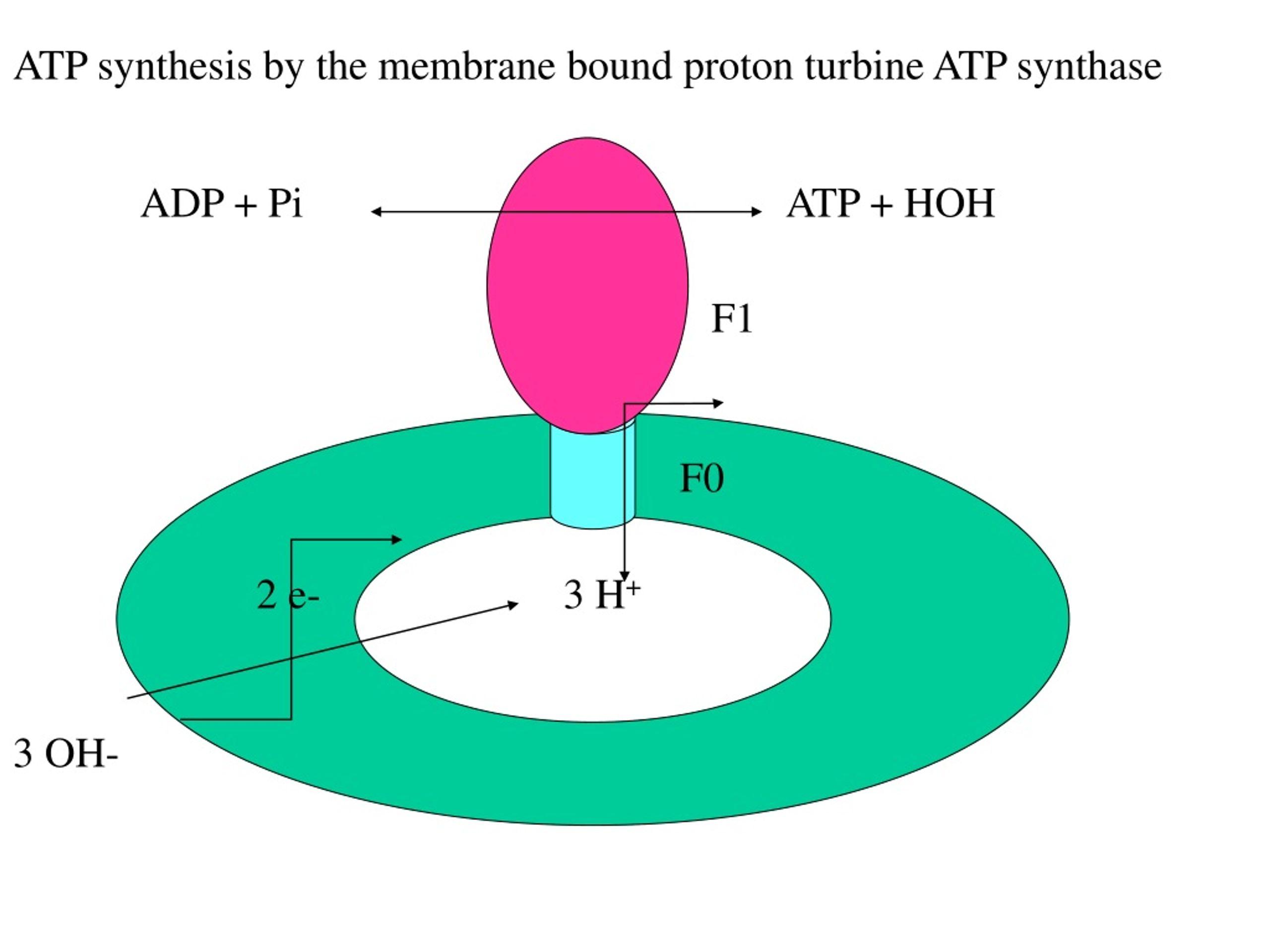
ATP synthesis occurs through the enzyme ATP synthase, which is embedded in the inner mitochondrial membrane.
- Protons flow back into the matrix through the F₀ portion of ATP synthase
- This flow causes the enzyme to rotate like a molecular turbine
- The rotational energy induces conformational changes in the F₁ portion
These structural changes enable the enzyme to bind ADP and inorganic phosphate (Pi) and convert them into ATP.
Energy Conversion in Chemiosmosis
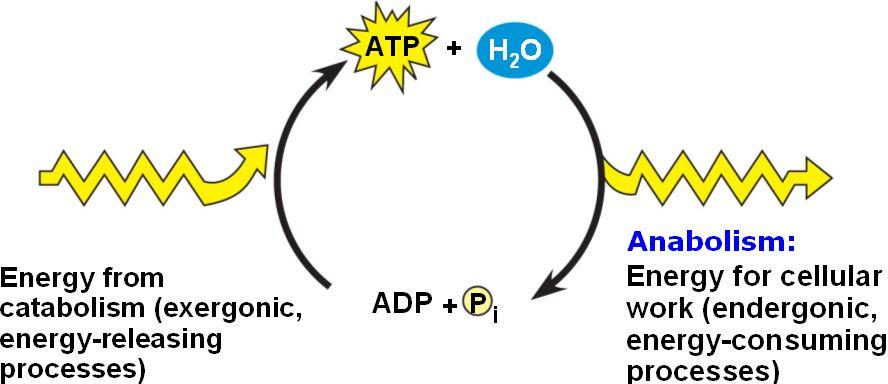
Chemiosmosis represents a highly efficient energy conversion process:
- Electrochemical energy (proton gradient) → converted into
- Mechanical energy (ATP synthase rotation) → which produces
- Chemical energy (ATP)
This step produces the majority of ATP in aerobic respiration, making oxidative phosphorylation the most energy-efficient pathway in living cells.
ATP Yield in Oxidative Phosphorylation
The ATP yield in Oxidative Phosphorylation represents the largest share of energy generated during cellular respiration. By utilizing electrons from NADH and FADH₂ in the Electron Transport Chain, cells efficiently convert biochemical energy into ATP.
ATP Produced per NADH and FADH₂

The number of ATP molecules generated depends on the source of electrons:
- NADH donates electrons at Complex I, leading to the pumping of more protons
→ Produces approximately 2.5 ATP per NADH - FADH₂ donates electrons at Complex II, bypassing Complex I
→ Produces approximately 1.5 ATP per FADH₂
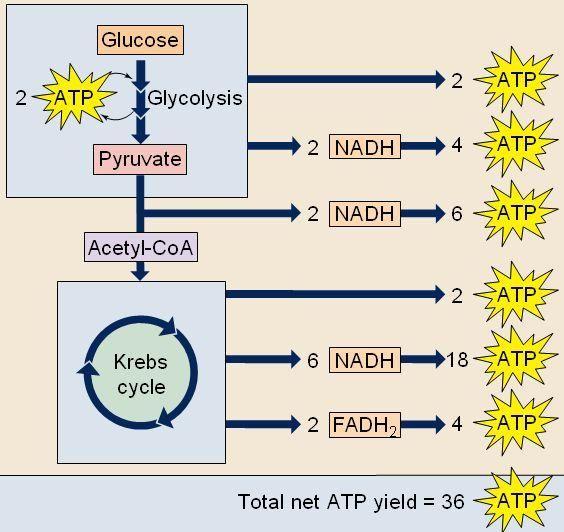
This difference occurs because FADH₂ contributes less to the proton gradient, resulting in lower ATP production.
Total ATP Produced in Oxidative Phosphorylation
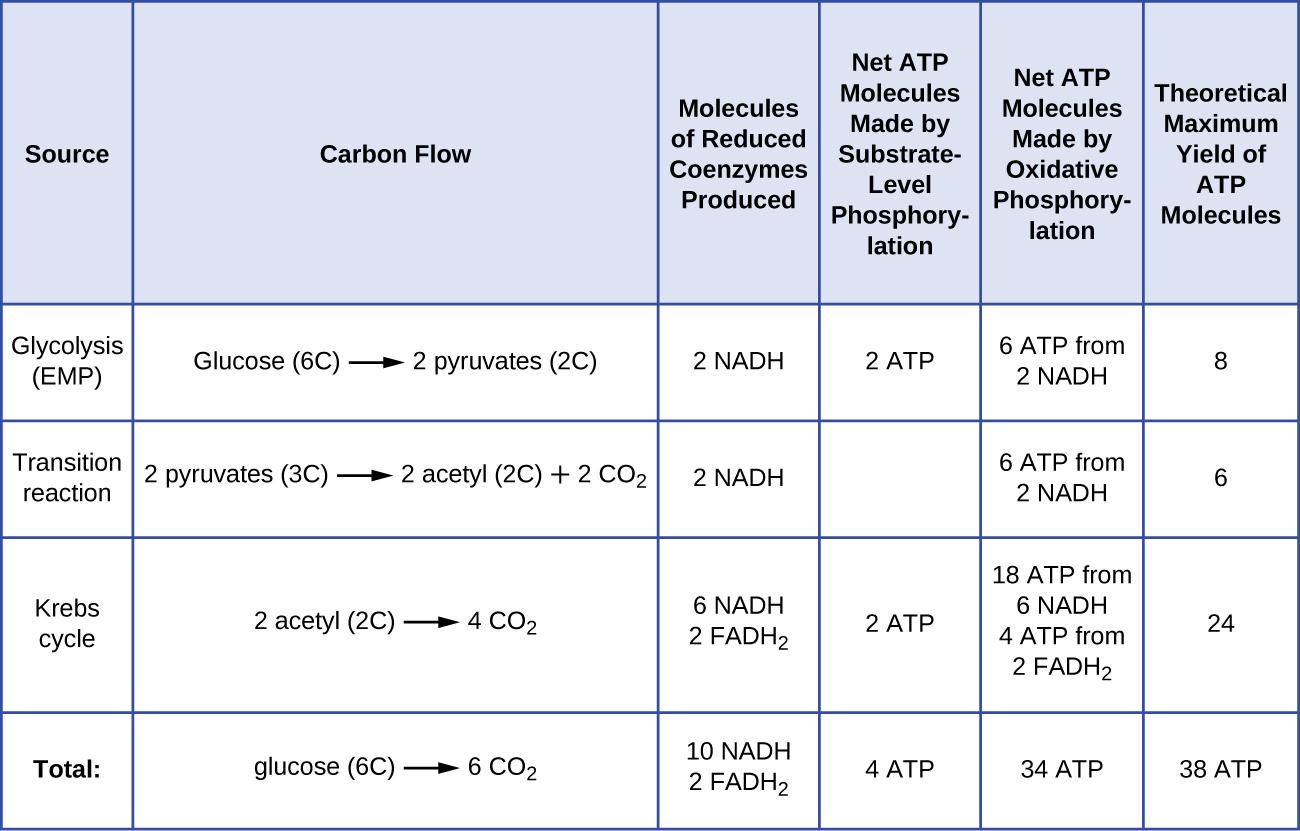
During complete aerobic respiration of one glucose molecule:
- NADH and FADH₂ generated from glycolysis and the Krebs Cycle feed into oxidative phosphorylation
- This results in the production of approximately 26–28 ATP molecules
This makes oxidative phosphorylation the most productive stage of cellular respiration, contributing the majority of ATP compared to earlier stages.
Theoretical vs Actual ATP Yield
The ATP yield in oxidative phosphorylation can vary between theoretical and actual values:
- Theoretical yield:
- Assumes perfect efficiency
- ~32 ATP per glucose (older estimate)
- Actual yield:
- Typically ~26–28 ATP per glucose
- Lower due to energy losses and cellular conditions
Reasons for this difference include:
- Proton leakage across the inner mitochondrial membrane
- Energy used for transporting molecules (e.g., ADP, Pi)
- Variations in shuttle systems (e.g., glycerol phosphate vs malate-aspartate shuttle)
Oxidative Phosphorylation vs Substrate-Level Phosphorylation
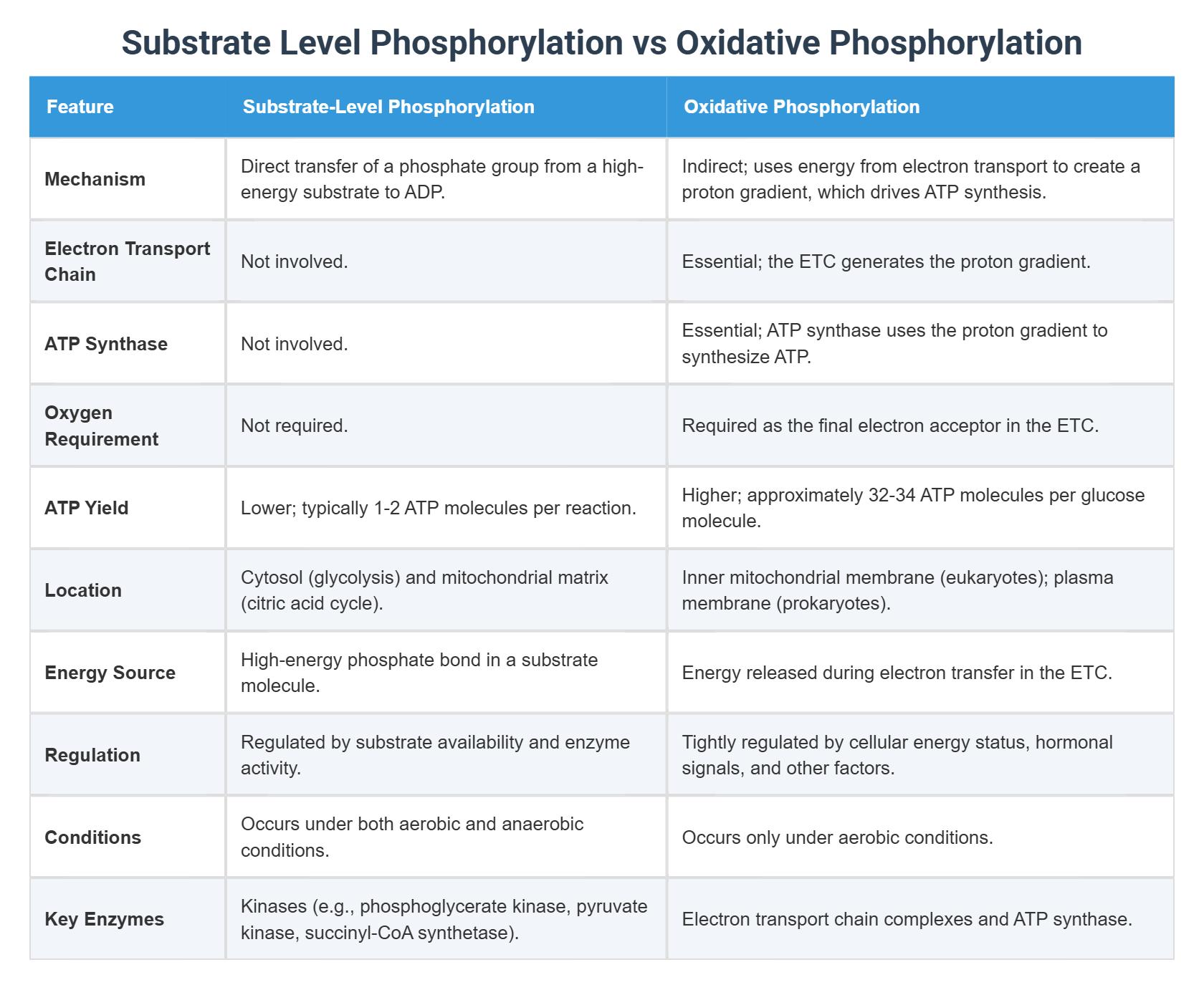
Understanding the difference between Oxidative Phosphorylation and substrate-level phosphorylation is essential for grasping how cells produce energy under different conditions.
| Feature | Oxidative Phosphorylation | Substrate-Level Phosphorylation |
|---|---|---|
| Location | Inner mitochondrial membrane | Cytoplasm / Mitochondria |
| Oxygen Requirement | Required (aerobic) | Not required (anaerobic possible) |
| ATP Yield | High (~26–28 ATP) | Low (2–4 ATP) |
| Mechanism | Proton gradient + chemiosmosis | Direct transfer of phosphate group |
Key Differences Explained
- Oxidative phosphorylation uses energy from electrons flowing through the Electron Transport Chain to create a proton gradient that drives ATP synthesis.
- Substrate-level phosphorylation directly transfers a phosphate group from a substrate molecule to ADP, forming ATP without the need for oxygen or an electron transport chain.
This makes oxidative phosphorylation far more efficient, while substrate-level phosphorylation acts as a backup energy source, especially during anaerobic conditions.
Inhibitors of Oxidative Phosphorylation
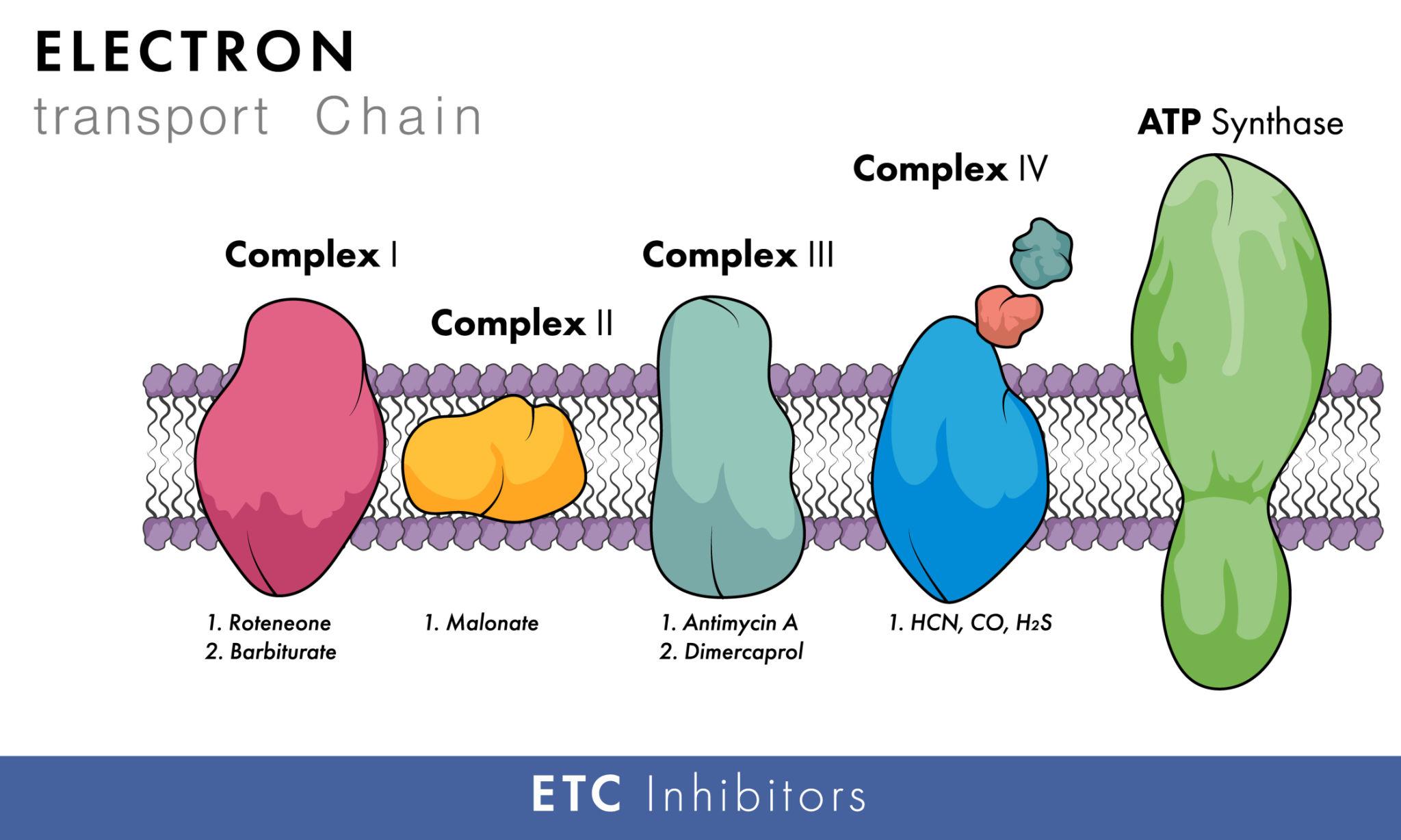
Certain chemicals can interfere with oxidative phosphorylation by blocking key steps in the electron transport chain or ATP synthesis.
Cyanide (ETC Inhibitor)
- Cyanide binds to Complex IV (cytochrome c oxidase)
- Prevents transfer of electrons to oxygen
- Stops the entire electron transport chain
👉 Result: No proton gradient formation → No ATP production → Rapid cell death
Oligomycin (ATP Synthase Inhibitor)
- Blocks the proton channel of ATP Synthase
- Prevents protons from flowing back into the matrix
- Halts ATP synthesis
👉 Result: Proton gradient builds up, but ATP cannot be produced
Effects on ATP Production
- Complete inhibition of ATP generation via oxidative phosphorylation
- Cells switch to anaerobic pathways (limited ATP supply)
- Can lead to severe cellular damage or death
These inhibitors highlight how critical each component of oxidative phosphorylation is for survival.
Regulation of Oxidative Phosphorylation
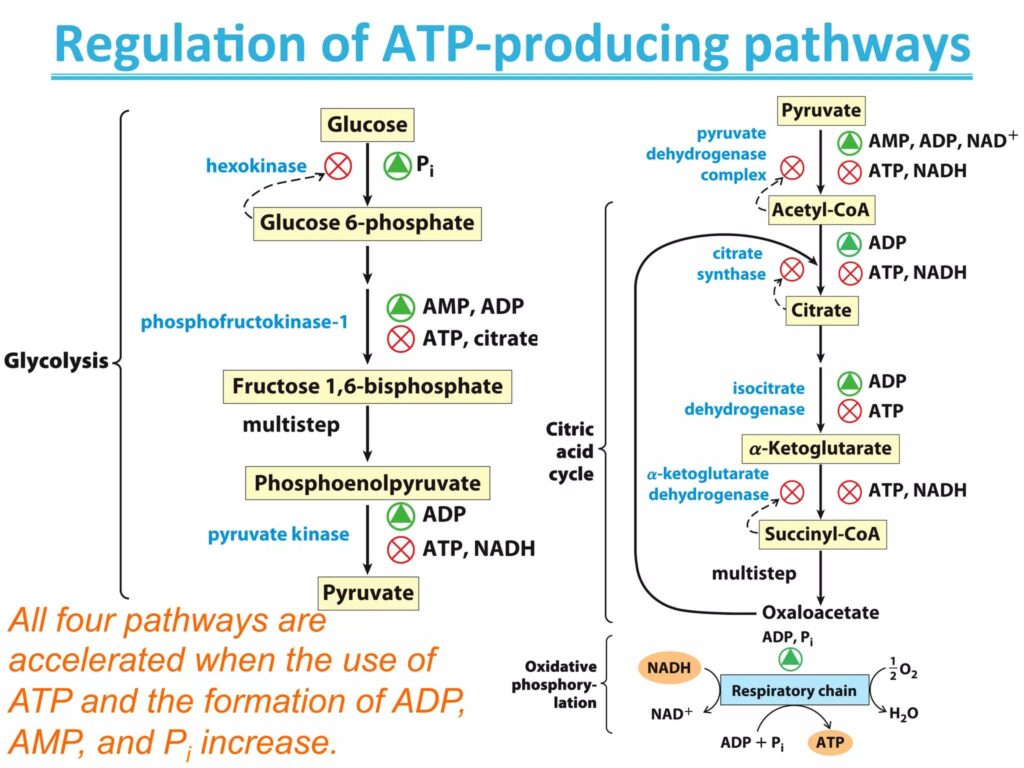
Oxidative phosphorylation is tightly regulated to match the energy needs of the cell. This ensures efficient ATP production without unnecessary energy waste.
Role of ADP/ATP Ratio
- High ADP levels signal low energy availability
- Stimulates electron transport chain activity and ATP synthesis
- High ATP levels slow down the process
👉 This balance ensures ATP is produced only when needed
Feedback Mechanisms
- Availability of NADH and FADH₂ controls electron flow
- Oxygen availability regulates the final step of the ETC
- Enzyme activity is adjusted based on cellular conditions
These feedback systems help maintain metabolic balance.
Energy Demand Control
- Cells increase oxidative phosphorylation during high energy demand (e.g., muscle activity)
- Decrease activity when energy demand is low
This dynamic control allows cells to adapt quickly to changing conditions and maintain optimal energy levels.
Hypotheses of Oxidative Phosphorylation
Several hypotheses have been proposed to explain how electron transport is coupled with ATP synthesis in Oxidative Phosphorylation. The three most important ones are discussed below:
1. Chemical Coupling Hypothesis
This hypothesis was proposed by Edward Slater and later supported by Albert Lehninger (1967).
According to this theory:
- Electron transport leads to the formation of high-energy chemical intermediates
- The breakdown of these intermediates drives ATP synthesis
Example: In glycolysis, oxidation of glyceraldehyde-3-phosphate (G3P) by NAD⁺ forms 1,3-bisphosphoglycerate, which donates a phosphate group to ADP to form ATP.
Limitation:
- No specific high-energy intermediate has been identified in mitochondria
- Due to lack of experimental evidence, this hypothesis is not widely accepted today
2. Conformational Coupling Hypothesis
This hypothesis was proposed by Paul D. Boyer.
According to this theory:
- Electron transport causes conformational (structural) changes in proteins of the inner mitochondrial membrane
- These structural changes are linked to ATP synthesis
- Energy stored in protein conformations drives ATP formation
Limitation:
- Limited experimental support
- Could not fully explain the mechanism of ATP generation
3. Chemiosmotic Hypothesis (Accepted Theory)
Peter Mitchell proposed the chemiosmotic theory in 1961 and was awarded the Nobel Prize in 1978.
This is the widely accepted explanation for oxidative phosphorylation.
Core Concept: Electron transport across the Electron Transport Chain generates a proton gradient across the inner mitochondrial membrane.
- Protons (H⁺) are pumped from the matrix to the intermembrane space
- This creates an electrochemical gradient (proton motive force, PMF)
- The PMF drives ATP synthesis via ATP Synthase
Experimental Evidence Supporting Chemiosmotic Hypothesis
- Electron transport creates a measurable proton gradient across the membrane
- The pH outside the membrane is lower than inside (difference ~1.4 units)
- Membrane potential (~0.14 V) is generated
- ATP can be synthesized artificially by creating a proton gradient (even without electron transport)
- Complexes I, III, and IV pump protons across the membrane
- Intact membrane structure is essential for ATP synthesis
Proton Pumping Mechanisms
Two mechanisms have been proposed to explain how protons are translocated:
(i) Redox Loop Mechanism
Proposed by Peter Mitchell, this mechanism is based on alternating carriers:
- Some carriers transfer hydrogen atoms (H)
- Others transfer electrons (e⁻)
This creates oxidation–reduction loops (O/R loops) in the respiratory chain.
- Each loop corresponds to coupling sites I, II, and III
- Protons are transported from matrix to intermembrane space
- Electrons cycle back through the membrane
👉 This contributes to the formation of the proton gradient required for ATP synthesis.
(ii) Proton Transport Mechanism
In this mechanism:
- Electron transport chain components are arranged to facilitate directional proton movement
- For every pair of electrons transferred from NADH to oxygen:
- Approximately 6 protons are pumped across the membrane (classical view)
This proton movement establishes the proton motive force that drives ATP synthesis.
Key Takeaway
- Early hypotheses (chemical and conformational) attempted to explain ATP formation but lacked strong evidence
- The chemiosmotic hypothesis is now universally accepted
- ATP synthesis is driven by a proton gradient, not a chemical intermediate
Summary of Oxidative Phosphorylation
Oxidative phosphorylation is the final and most efficient stage of cellular respiration, responsible for generating the majority of ATP in aerobic organisms. It integrates the Electron Transport Chain with chemiosmosis to convert energy stored in electron carriers into usable cellular energy.
Bullet Point Recap
- Occurs in the inner mitochondrial membrane
- Involves the transfer of electrons from NADH and FADH₂ through the electron transport chain
- Energy released during electron transfer is used to pump protons (H⁺) across the membrane
- Creates a proton gradient (proton motive force)
- Protons flow back through ATP Synthase
- Drives the synthesis of ATP from ADP and inorganic phosphate
- Oxygen acts as the final electron acceptor, forming water
Key Takeaways
- Oxidative phosphorylation produces ~26–28 ATP per glucose, making it the most productive stage of cellular respiration
- It depends on oxygen and a functional electron transport chain
- The proton gradient is the central energy source driving ATP synthesis
- Any disruption in this process can significantly impact cellular energy production
Reinforcing the Main Concept
At its core, oxidative phosphorylation is a highly efficient energy conversion system. It transforms the energy carried by electrons into a proton gradient, which is then used to power ATP synthesis. This elegant mechanism ensures that cells can meet their energy demands and sustain life processes effectively.
Important Points for Exams
These quick points are essential for exams and rapid revision:
- Location: Inner mitochondrial membrane
- Process Type: Aerobic (requires oxygen)
- Final Electron Acceptor: Oxygen (O₂)
- Main Output: ATP (majority of cellular energy)
- Mechanism: Driven by proton gradient (chemiosmosis)
- Key Components: Electron transport chain + ATP synthase
- Byproduct: Water (H₂O)
Exam Tip: Focus on the sequence.
Electron Transport Chain → Proton Gradient → ATP Synthase → ATP Production
Remembering this flow helps you quickly answer both conceptual and diagram-based questions.
FAQs on Oxidative Phosphorylation
What is the main function of oxidative phosphorylation?
The main function of oxidative phosphorylation is to produce ATP by using energy released from electrons passing through the Electron Transport Chain. This process generates a proton gradient that drives ATP synthesis through chemiosmosis.
How much ATP is produced in oxidative phosphorylation?
Oxidative phosphorylation produces approximately 26–28 ATP molecules per glucose. The exact number may vary depending on cellular conditions and the efficiency of the electron transport chain.
Why is oxygen required in oxidative phosphorylation?
Oxygen is required because it acts as the final electron acceptor in the electron transport chain. It accepts electrons and combines with protons to form water, allowing the process to continue efficiently.
What happens if oxygen is absent?
In the absence of oxygen, the electron transport chain stops functioning, and oxidative phosphorylation cannot occur. As a result, ATP production decreases significantly, and cells switch to Anaerobic Respiration for limited energy production.
Where does oxidative phosphorylation occur?
Oxidative phosphorylation occurs in the inner mitochondrial membrane, where the electron transport chain and ATP Synthase are located.
What is chemiosmosis in oxidative phosphorylation?
Chemiosmosis is the process by which ATP is generated using the energy of a proton gradient. Protons flow through ATP synthase, driving the conversion of ADP into ATP.
What is the role of NADH and FADH₂?
NADH and FADH₂ act as electron carriers. They donate high-energy electrons to the electron transport chain, which drives proton pumping and ATP production.
What are the main components of oxidative phosphorylation?
The main components include:
– Electron transport chain (Complex I–IV)
– Proton gradient (proton motive force)
– ATP synthase
Final words on Oxidative Phosporylation
Oxidative phosphorylation is not just another step in cellular respiration—it is the powerhouse mechanism that fuels life at the molecular level. By efficiently converting energy from electrons into ATP through the electron transport chain and chemiosmosis, this process ensures that cells have a continuous supply of energy to perform essential functions.
From understanding the role of oxygen to exploring ATP yield and regulatory mechanisms, mastering oxidative phosphorylation gives you a complete picture of how energy flows within living systems. Whether you’re a student preparing for exams or a learner building strong fundamentals in biochemistry, this topic forms a critical foundation.
What to Do Next?
- Bookmark this guide for quick revision before exams
- Explore related topics like Glycolysis and Krebs Cycle to complete your understanding of cellular respiration
- Have a question? Drop it in the comments and keep the discussion going
- Found this helpful? Share it with your classmates or fellow learners
In One Line: Oxidative phosphorylation is the process by which cells use oxygen and electron transport to generate a proton gradient that drives large-scale ATP production.
Discover more from Biochemistry Den
Subscribe to get the latest posts sent to your email.