Bonds in Protein Structure: A Complete Guide for Students
Proteins are the workhorses of every living cell — they catalyze reactions, provide structural support, transport molecules, and defend the body against disease. But none of this is possible without one thing: bonds.
The bonds in protein structure determine its shape, and shape determines function. Diverse essential molecular processes within a cell are carried out by molecular machines built from a large number of protein components organized by their Protein-Proteins interaction (PPI).
Indeed, these interactions are at the core of the entire interactomics system of any living cell. So, unsurprisingly, aberrant PPIs are based on multiple diseases, such as Creutzfeld-Jacob, Alzheimer’s disease, and cancer.
Whether you’re a medical student, a biology undergraduate, or preparing for the MCAT/NEET, this guide covers everything you need to know about bonds in protein structure—clearly, completely, and in one place.
Why Are Bonds Important in Protein Structure?
A protein is not just a random chain of amino acids. It folds into a precise three-dimensional shape that is critical for its biological function. This folding and the stability of that final shape depend entirely on the chemical bonds and interactions between amino acids.
Without these bonds:
- Proteins would collapse into non-functional random coils
- Enzymes would lose their active sites and stop catalyzing reactions
- Structural proteins like collagen and keratin would fail to provide mechanical support
- Antibodies could not bind to antigens with specificity
In short, bonds in protein structure are what make life possible. There are two broad categories: covalent bonds (strong, permanent) and non-covalent interactions (weaker, but collectively powerful and reversible). Together they sculpt the protein from a linear sequence into a functional 3D molecule.
Types of Bonds in Protein Structure
Proteins use not one, but six distinct types of bonds and interactions to build and maintain their structure. These range from strong covalent bonds that form the protein backbone to weaker non-covalent interactions that guide folding and determine the final 3D shape.
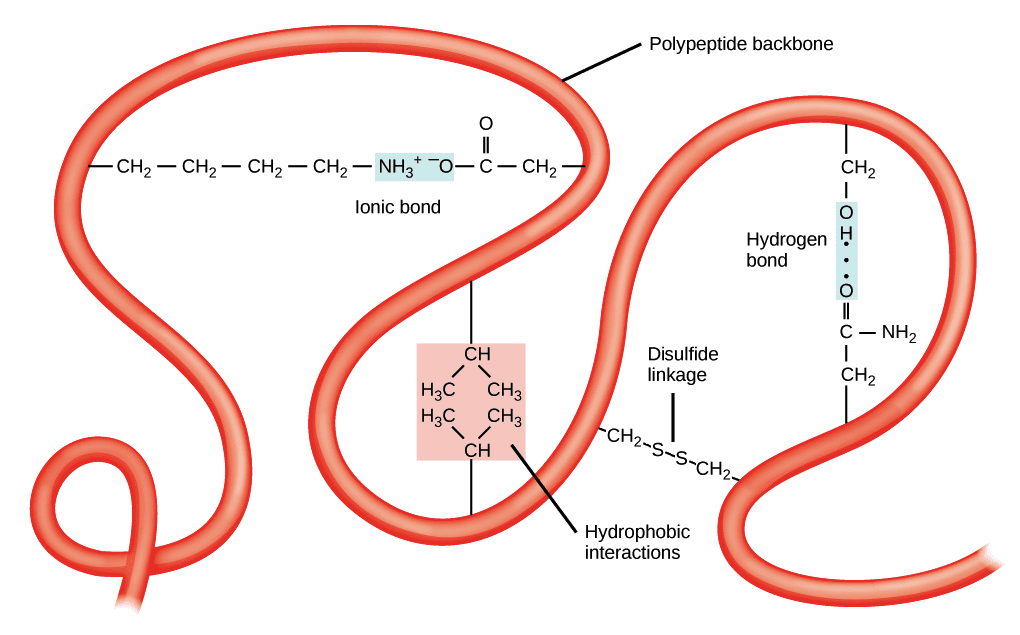
Understanding each bond type — how it forms, how strong it is, and where it appears — is essential for grasping how proteins achieve their remarkable structural diversity and functional specificity.
Let’s explore each one in detail.
1. Peptide Bonds (Primary Structure)
The peptide bond is the backbone bond of all proteins. It forms between the carboxyl group (–COOH) of one amino acid and the amino group (–NH₂) of the next, with the release of one water molecule—a reaction called dehydration synthesis (condensation reaction).
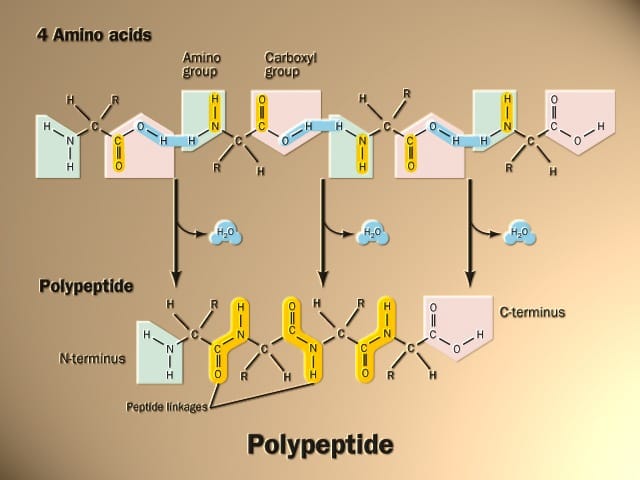
Key features of peptide bonds:
- They are covalent bonds—the strongest type in protein chemistry
- Bond energy is approximately 200–400 kJ/mol
- They have partial double-bond character due to resonance, making the bond rigid and planar
- The C–N peptide bond does not rotate freely, which restricts the backbone conformation
- The resulting chain is called a polypeptide
Example: When glycine and alanine join, the carboxyl of glycine reacts with the amino of alanine, releasing water and forming Gly-Ala with a peptide bond between them.
💡 Student Tip: The peptide bond is always written as –CO–NH–. The carbon belongs to the preceding amino acid; the nitrogen belongs to the next.
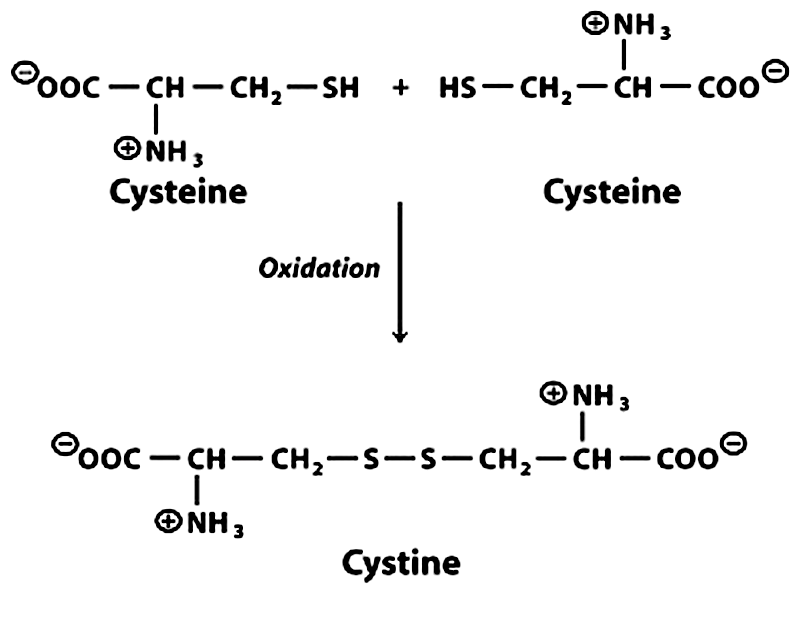
2. Disulfide Bonds (Tertiary Structure)
Disulfide bonds are strong covalent bonds formed between the sulfhydryl groups (–SH) of two cysteine amino acid residues. The reaction involves oxidation—two –SH groups lose hydrogen atoms to form an –S–S– (disulfide bridge).
Key features:
- Bond energy: ~167 kJ/mol
- Only formed by cysteine residues
- Critical for tertiary and quaternary structure stabilization
- Found in extracellular proteins (e.g., antibodies, insulin) where the oxidizing environment supports their formation
- Intracellular proteins rarely have disulfide bonds because the cytoplasm is reducing
Classic examples:
- Insulin: Contains 2 inter-chain and 1 intra-chain disulfide bonds that link its A and B chains
- Immunoglobulins (antibodies): Multiple disulfide bonds stabilize the heavy and light chains
- Keratin: Rich in disulfide bonds—this is why hair can be permanently waved (bonds broken and reformed in new position)
💡 Student Tip: Disulfide bonds are broken by reducing agents like β-mercaptoethanol or dithiothreitol (DTT) — a key fact for lab and exam questions.
3. Hydrogen Bonds (Secondary & Tertiary Structure)
Hydrogen bonds are non-covalent interactions that form when a hydrogen atom covalently bonded to an electronegative atom (like N or O) is attracted to another electronegative atom nearby.
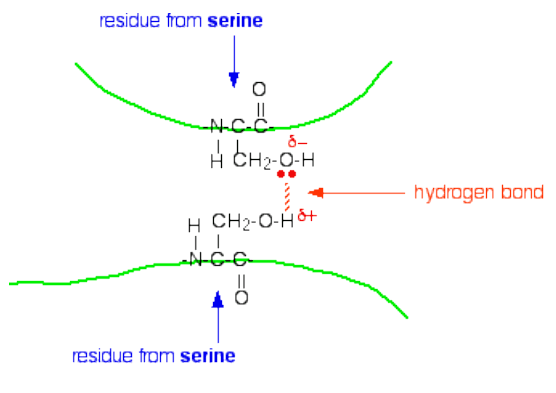
In proteins, hydrogen bonds form primarily between:
- The backbone –C=O (hydrogen bond acceptor) and –NH (hydrogen bond donor) groups of the polypeptide chain
Key features:
- Bond energy: ~5–30 kJ/mol (individually weak, but collectively very strong)
- They are the primary force stabilizing secondary structure (α-helix and β-sheet)
- Also contribute to tertiary structure via side-chain hydrogen bonding
- They are reversible—easily broken and reformed
In the α-helix: Every backbone –NH donates a hydrogen bond to the –C=O of the amino acid 4 residues earlier in the chain. This creates a right-handed helical coil stabilized by hydrogen bonds running parallel to the helix axis.
In the β-pleated sheet: Hydrogen bonds form between adjacent polypeptide strands (parallel or antiparallel), creating a sheet-like structure. Antiparallel β-sheets have stronger, more linear hydrogen bonds and are more stable.
💡 Student Tip: Hydrogen bonds do NOT involve covalent sharing of electrons. They are purely electrostatic attractions—which is why they break at high temperatures (denaturation).
4. Ionic Bonds (Salt Bridges)
Ionic bonds in proteins — also called salt bridges or electrostatic interactions — form between oppositely charged amino acid side chains.
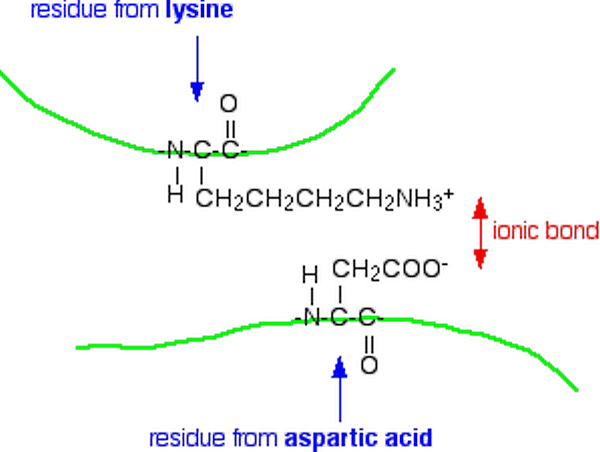
Positively charged (at physiological pH ~7.4):
- Lysine (Lys, K) — –NH₃⁺
- Arginine (Arg, R) — guanidinium group
- Histidine (His, H) — imidazolium group
Negatively charged (at physiological pH):
- Aspartate (Asp, D) — –COO⁻
- Glutamate (Glu, E) — –COO⁻
Key features:
- Bond energy: ~20 kJ/mol
- pH-dependent—changing pH neutralizes charges and disrupts these bonds (one reason acids/bases denature proteins)
- Salt bridges are often found on the protein surface where they interact with the aqueous environment
- They contribute primarily to tertiary and quaternary structure
Example: A lysine residue (–NH₃⁺) on one part of the chain forms an ionic interaction with a glutamate residue (–COO⁻) on another part, pulling the two regions together.
5. Van der Waals Forces
Van der Waals forces are weak, transient, non-covalent interactions that arise from temporary fluctuations in electron distribution around atoms, creating instantaneous dipoles.
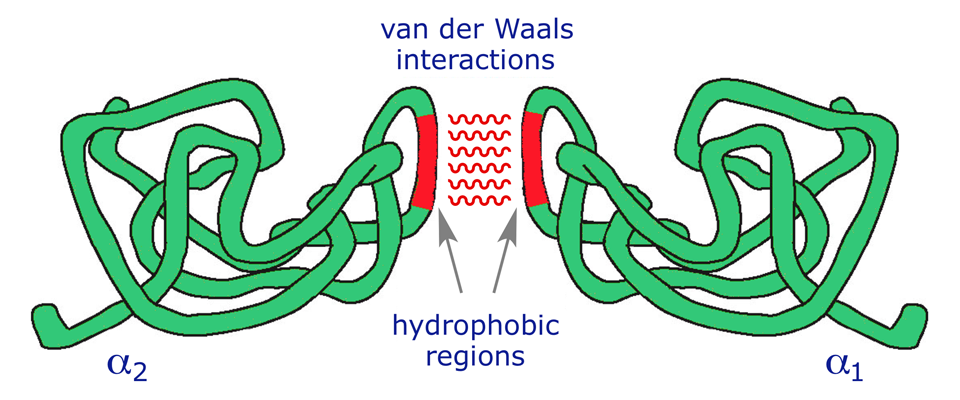
Key features:
- Bond energy: ~0.4–4 kJ/mol (extremely weak individually)
- Operate only over very short distances (~0.3–0.5 nm)
- Significant in the hydrophobic core of proteins, where non-polar side chains are tightly packed
- They are additive — thousands of Van der Waals contacts in a protein collectively contribute significantly to stability
Why they matter: Although each Van der Waals interaction is trivially weak, a tightly packed protein interior can have thousands of them. Their collective contribution to folding stability is substantial and should not be underestimated.
💡 Student Tip: Van der Waals forces exist between ALL atoms in proximity, but they are most functionally relevant in the protein’s hydrophobic interior where other stronger interactions are absent.
6. Hydrophobic Interactions
Hydrophobic interactions are arguably the most important driving force for protein folding. They are not true bonds — there is no attractive force between nonpolar groups — but rather an entropy-driven phenomenon.
How they work:
When a protein folds in aqueous solution, nonpolar (hydrophobic) side chains (e.g., valine, leucine, isoleucine, phenylalanine) are driven into the interior of the protein away from water. This maximizes the entropy of the surrounding water molecules (water no longer has to form ordered cages around hydrophobic groups).
Key features:
- Not a true bond — no electron sharing or charge attraction
- Driven by thermodynamics (increase in entropy of water)
- Major contributor to tertiary and quaternary structure
- Disrupted by detergents and urea (which solubilize hydrophobic groups)
Amino acids involved: Val, Leu, Ile, Met, Phe, Trp, Pro, Ala (nonpolar side chains)
Bonds at Each Level of Protein Structure
Proteins are organized into four hierarchical levels of structure—primary, secondary, tertiary, and quaternary—and each level is governed by a distinct set of bonds and interactions.
As you move up from the simple amino acid chain to the fully folded, multi-subunit protein, the variety and complexity of bonding increases.
Knowing which bonds operate at which structural level is one of the most commonly tested concepts in biochemistry exams worldwide—and one of the most important for understanding how proteins function in living cells.
1. Primary Structure Bonds
The primary structure is the linear sequence of amino acids in a polypeptide chain. It is held together exclusively by peptide bonds—covalent bonds formed during ribosomal translation. The sequence of amino acids is encoded by DNA and is unique to each protein. The primary structure ultimately determines all higher levels of structure because the amino acid sequence dictates how the protein folds.
2. Secondary Structure Bonds
Secondary structure refers to locally ordered, repetitive folding patterns within the polypeptide chain. The two most common are:
- Alpha-helix (α-helix): Stabilized entirely by hydrogen bonds between backbone -NH and -C=O groups 4 residues apart. Proline cannot participate (no -NH group) and acts as a “helix breaker.”
- Beta-pleated sheet (β-sheet): Stabilized by hydrogen bonds between backbone groups of adjacent strands. Can be parallel or antiparallel.
- Beta-turns and loops: Stabilized by hydrogen bonds and sometimes ionic interactions; connect helices and strands. The dominant bond in secondary structure is the hydrogen bond.
3. Tertiary Structure Bonds
Tertiary structure is the overall 3D shape of a single polypeptide chain—the final folded conformation. It is stabilized by interactions between amino acid side chains (R groups), including:
- Disulfide bonds (covalent — strongest tertiary interaction)
- Hydrogen bonds between polar side chains
- Ionic bonds/salt bridges between charged side chains
- Hydrophobic interactions (major driving force)
- Van der Waals forces in the packed interior
The combination of all these interactions determines the protein’s unique 3D shape and, therefore, its function.
4. Quaternary Structure Bonds
Quaternary structure exists only in proteins made of two or more polypeptide subunits (protomers). The subunits are held together by:
- Hydrogen bonds between subunit surfaces
- Ionic bonds/salt bridges at subunit interfaces
- Hydrophobic interactions between nonpolar patches on subunit surfaces
- Van der Waals forces
- Occasionally disulfide bonds between subunits (e.g., immunoglobulins)
Classic example: Hemoglobin — a tetramer of 2 α-subunits and 2 β-subunits held together by non-covalent interactions. When oxygen binds to one subunit, it causes conformational changes that affect the other subunits—a process called “cooperative binding” made possible by these inter-subunit bonds.
Comparison Table: Covalent vs Non-Covalent Bonds
With six types of bonds involved in protein structure, it can be challenging to keep track of their differences.
The most fundamental distinction is between covalent bonds—which involve electron sharing and are strong and permanent—and non-covalent interactions—which are individually weaker but reversible and collectively powerful.
The table below summarizes all six bond types side by side, so you can quickly compare their strength, location, reversibility, and the amino acids involved, making revision faster and more effective.
| Bond Type | Category | Strength (kJ/mol) | Structure Level | Reversible? | Key Amino Acids |
|---|---|---|---|---|---|
| Peptide Bond | Covalent | 200–400 | Primary | ❌ No | All amino acids |
| Disulfide Bond | Covalent | ~167 | Tertiary / Quaternary | ❌ No* | Cysteine only |
| Hydrogen Bond | Non-covalent | 5–30 | Secondary & Tertiary | ✅ Yes | Ser, Thr, Asn, Gln, backbone |
| Ionic Bond (Salt Bridge) | Non-covalent | ~20 | Tertiary / Quaternary | ✅ Yes | Lys, Arg, His, Asp, Glu |
| Van der Waals | Non-covalent | 0.4–4 | Tertiary | ✅ Yes | All nonpolar residues |
| Hydrophobic Interaction | Non-covalent | Variable | Tertiary / Quaternary | ✅ Yes | Val, Leu, Ile, Phe, Trp |
*Disulfide bonds can be reduced/re-oxidized under specific conditions.
Key Insight: Covalent bonds (peptide and disulfide) provide a permanent structural backbone, while non-covalent interactions provide flexibility, adaptability, and reversibility—essential for functions like enzyme catalysis and protein-ligand binding.
Clinical Significance: Disease & Denaturation
Understanding bonds in protein structure isn’t just academic—it has real medical implications.
Protein Denaturation
Denaturation is the disruption of a protein’s 3D structure (secondary, tertiary, quaternary) without breaking peptide bonds. The primary structure (amino acid sequence) remains intact, but the protein loses its shape and function.
What breaks each bond type during denaturation:
| Denaturing Agent | Bonds Disrupted |
|---|---|
| Heat (high temperature) | Hydrogen bonds, hydrophobic interactions, ionic bonds |
| Acids / Bases | Ionic bonds (by changing charge state) |
| Urea / Guanidinium chloride | Hydrogen bonds, hydrophobic interactions |
| Detergents (SDS) | Hydrophobic interactions |
| Reducing agents (β-mercaptoethanol, DTT) | Disulfide bonds |
| Organic solvents | Hydrophobic interactions, hydrogen bonds |
Note: Denaturation is often irreversible in biological systems (e.g., cooked egg white), though some small proteins can renature spontaneously when the denaturing agent is removed (Anfinsen’s experiment with ribonuclease A—a Nobel Prize-winning demonstration that the amino acid sequence alone contains all information needed for folding).
Disease Connections
- Sickle Cell Anemia: A single amino acid substitution (Glu→Val at position 6 of the β-globin chain) changes a charged, hydrophilic surface residue to a hydrophobic one. This creates abnormal hydrophobic interactions between hemoglobin molecules, causing them to polymerize into rigid fibers—resulting in the sickle shape of red blood cells.
- Alzheimer’s Disease: Misfolding of tau proteins and amyloid-beta peptides leads to pathological aggregation. Amyloid-beta forms abnormal β-sheet-rich fibrils stabilized by intermolecular hydrogen bonds—forming the plaques seen in Alzheimer’s disease.
- Creutzfeldt-Jakob Disease (Prion Diseases): Prion proteins misfold from their normal α-helix-rich (PrPᶜ) form into a β-sheet-rich pathological form (PrPˢᶜ). The abnormal form recruits normal proteins to misfold—driven by altered hydrogen bonding patterns.
- Cystinuria: A genetic disorder where cystine (oxidized cysteine dimer with a disulfide bond) accumulates in urine, forming kidney stones—a direct consequence of disulfide bond chemistry.
- Osteogenesis Imperfecta (Brittle Bone Disease): Mutations in collagen disrupt the triple-helix structure of collagen, which is stabilized by hydrogen bonds and requires glycine at every third position. A single glycine substitution destabilizes the entire helix.
Frequently Asked Questions (FAQ)
What is the strongest bond in protein structure?
The peptide bond is the strongest bond in protein structure, with a bond energy of 200–400 kJ/mol. It is a covalent bond that forms the backbone of all polypeptide chains and is not broken except by proteolytic enzymes or harsh chemical conditions.
Which bonds are responsible for secondary structure?
Hydrogen bonds between backbone –NH and –C=O groups are the primary bonds responsible for secondary structure — including the α-helix and β-pleated sheet. These bonds do not involve amino acid side chains.
What bonds are broken during protein denaturation?
Denaturation breaks non-covalent bonds—hydrogen bonds, ionic bonds, hydrophobic interactions, and Van der Waals forces—disrupting secondary, tertiary, and quaternary structure. Peptide bonds are NOT broken during denaturation (the primary structure remains intact). Disulfide bonds may or may not be broken depending on the denaturing agent (reducing agents are needed to break S–S bonds).
How many types of bonds are in proteins?
There are 6 major types of bonds and interactions in protein structure: (1) peptide bonds, (2) disulfide bonds, (3) hydrogen bonds, (4) ionic bonds/salt bridges, (5) Van der Waals forces, and (6) hydrophobic interactions.
What is the difference between covalent and non-covalent bonds in proteins?
Covalent bonds (peptide and disulfide bonds) involve the sharing of electrons between atoms and are strong (100–400 kJ/mol), permanent, and not easily reversible. Non-covalent interactions (hydrogen bonds, ionic bonds, Van der Waals, hydrophobic) do not involve electron sharing, are individually weaker (0.4–30 kJ/mol), and are reversible — but collectively they are the primary determinants of 3D protein shape.
Why are non-covalent bonds important if they are so weak?
Though individually weak, non-covalent bonds are present in thousands of copies throughout a protein. Their collective effect creates a stable but dynamic structure. Crucially, their reversibility allows proteins to change shape (conformational changes) during enzyme catalysis, signal transduction, and molecular recognition — none of which would be possible if proteins were rigid covalently-locked structures.
Which bond is unique to proteins and not found in other biomolecules?
The peptide bond is unique to proteins (and peptides). While disulfide bonds are also found in some other biomolecules, the peptide bond—linking amino acids via –CO–NH– — is the defining bond of protein primary structure.
What is the role of proline in protein structure?
Proline is a unique amino acid with a cyclic side chain that bonds to its own backbone nitrogen, eliminating the –NH group. This means proline cannot donate hydrogen bonds and introduces a rigid kink in the polypeptide chain, making it a “helix breaker.” It is commonly found in turns and loops.
Summary: Bonds in Protein Structure at a Glance
Bonds in protein structure are not just a chemistry topic — they are the molecular foundation of life itself. From the rigid peptide bond that strings amino acids together to the thousands of weak non-covalent interactions that collectively fold a protein into its precise functional shape, every bond plays a defined role at a specific structural level.
Here is a quick recap of the key takeaways:
- Peptide bonds form the covalent backbone of every protein (primary structure) and are the strongest bonds present
- Disulfide bonds lock tertiary and quaternary structures in place, especially in extracellular and secreted proteins like insulin and antibodies
- Hydrogen bonds are the architects of secondary structure—they build every α-helix and β-sheet with clockwork precision
- Ionic bonds (salt bridges) form between oppositely charged side chains and are sensitive to changes in pH
- Van der Waals forces are individually negligible but collectively significant, especially in the tightly packed hydrophobic core
- Hydrophobic interactions are the single most important driving force for protein folding, governed by thermodynamics rather than direct attraction
- Denaturation disrupts non-covalent bonds and unfolds proteins—while the primary sequence survives, function is lost
- Diseases like sickle cell anemia, Alzheimer’s, and prion diseases all arise from disruptions in normal bonding and folding patterns
A helpful way to remember the hierarchy: covalent bonds build the scaffold; non-covalent interactions sculpt the architecture. Neither is sufficient alone — it is the interplay of all six bond types that produces a biologically active protein.
As you study protein biochemistry further, you will find that understanding these bonds is the key to understanding enzyme mechanisms, drug design, protein engineering, and the molecular basis of disease. Every time a drug binds its target, every time an enzyme catalyzes a reaction, and every time a muscle contracts, bonds in protein structure are at work.
💡 Quick Memory Aid:
“Pretty Disgusting Homework Is Very Hard”
→ Peptide · Disulfide · Hydrogen · Ionic · Van der Waals · Hydrophobic
Discover more from Biochemistry Den
Subscribe to get the latest posts sent to your email.





