What is Gluconeogenesis? What are the steps and importance of metabolism?
Gluconeogenesis is the process of synthesizing glucose from non-carbohydrate sources. Where does gluconeogenesis occur? The process takes place mainly in the liver and to a limited extent in the kidney and small intestine under some conditions.
It is also called “endogenous glucose production” (EGP). It is one of the metabolic pathways.
The production of glucose from other carbon skeletons is necessary since the testes, erythrocytes, and kidney medulla exclusively utilize glucose for ATP production.

Why is gluconeogenesis important? It ensures a continuous supply of glucose to glucose-dependent tissues such as the brain, erythrocytes, and renal medulla, especially during fasting, prolonged exercise, or low-carbohydrate intake.
Gluconeogenesis definition
The definition of Gluconeogenesis is given below
The biosynthesis of carbohydrates from simpler, non-carbohydrate precursors such as oxaloacetate and pyruvate is called “gluconeogenesis“.
(or)
“Synthesis of Glucose / Carbohydrates from Non-carbohydrate Precursor Molecules.”
- The starting point precursor of the pathways is a pyruvic acid molecule, although oxaloacetic acid and dihydroxyacetone phosphate also provide entry points.
- Lactic acid — a major gluconeogenic precursor — comes primarily from anaerobic glycolysis in muscle and erythrocytes, while glycerol is derived from the hydrolysis of triacylglycerols in adipose tissue.
- It is similar but not the exact reverse of glycolysis; some steps are identical in the reverse direction, and three of them are new ones.
- Glycolysis and gluconeogenesis are both reversible processes.
- Without going into detail, the general neoglucogenesis sequence is given in the graphic on the left.
- Notice that oxaloacetic acid is synthesized from pyruvic acid in the first step. Oxaloacetic acid is also the first compound to react with acetyl CoA in the citric acid cycle. The concentration of acetyl CoA and ATP determines the fate of oxaloacetic acid.
- If the concentration of acetyl CoA is low and the concentration of ATP is high, then neoglucogenesis proceeds. Also, notice that ATP is required for the biosynthesis sequence of the pathway.
- Gluconeogenesis occurs mainly in the liver, with a small amount also occurring in the cortex of the kidney. It occurs very little in the brain, skeletal muscles, heart muscles, or other body tissue. In fact, these organs have a high demand for glucose.
- Therefore, this pathway is constantly occurring in the liver to maintain the glucose level in the blood to meet these demands.
Steps in Gluconeogenesis
The synthesis of glucose from pyruvate utilizes many of the same enzymes as Glycolysis. Here is the gluconeogenesis pathway.
Kreb’s pointed out that energy barriers obstruct a simple reversal of glycolysis.
- Between Pyruvate and PEP (Enzymes: Pyruvate Carboxylase and Phosphoenolpyruvate Carboxylase-PEPCK)
- Between Fructose-1,6-bisP and Fructose-6-P (Enzymes: Enzyme: Fructose-1,6-bisphosphatase)
- Between Glucose-6-P and Glucose (Enzymes: Glucose-6-Phosphatase)
- Between Glucose-1-P and Glycogen (Enzyme: Glycogen Synthase)
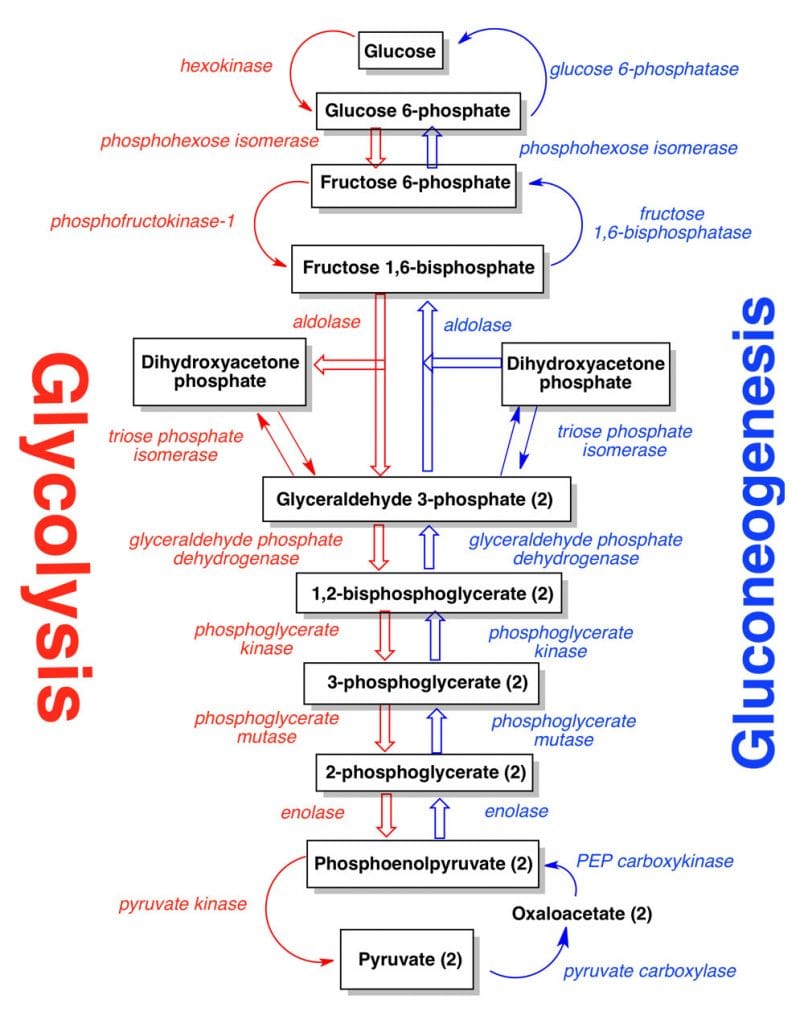
Glycolysis has three reactions that have a forward direction in that they are essentially irreversible (see lecture notes on Glycolysis):
- Hexokinase (or glucokinase)
- Phosphofructokinase, and
- Pyruvate Kinase.
These steps must be bypassed in gluconeogenesis. Two of the bypass reactions involve simple hydrolysis reactions.
Below is the forward reaction catalyzed by each of these glycolysis enzymes, followed by the bypass reaction catalyzed by the Gluconeogenesis enzyme.
Step 1: Glucose Phosphorylation/Dephosphorylation
In Glycolysis, the first step is phosphorylation.
Glucose + ATP –> Glucose-6-phosphate + ADP
Enzyme: Hexokinase or Glucokinase (Glycolysis)
In gluconeogenesis, the first step in glycolysis is reversible.
Glucose-6-phosphate + H₂O –> glucose + Pi
Enzyme: Glucose-6-phosphatase
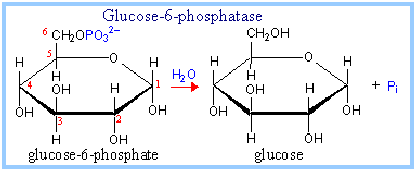
The glucose-6-phosphatase enzyme is embedded in the endoplasmic reticulum (ER) membrane in the liver of the cells.
Evidence indicates that the catalytic site is exposed to the ER lumen. Another subunit of the enzyme is postulated to function as a translocase, providing substrate access to the active site.
Step 2: Fructose Phosphorylation/Dephosphorylation
In Glycolysis, Step 3 is
Fructose-6-phosphate + ATP –> fructose-1,6-bisphosphate + ADP
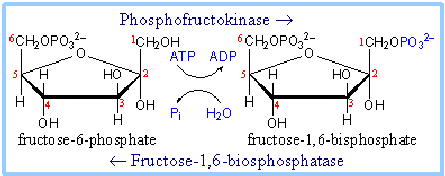
Enzymes: Phosphofructokinase
In gluconeogenesis, step 3 in glycolysis is reversible.
Fructose-1,6-bisphosphate + H₂O –> fructose-6-phosphate + Pi
Enzyme: Fructose-1,6-Bisphosphatase
Step 3: Pyruvate Phosphorylation/Dephosphorylation
In glycolysis, step 9 is
Phosphoenolpyruvate + ADP –> pyruvate + ATP
Enzyme: Pyruvate Kinase
For bypass of the pyruvate kinase reaction of glycolysis, cleavage of ~2 P bonds is required. The free energy change associated with the cleavage of one P bond of ATP is insufficient to drive the synthesis of phosphoenolpyruvate (PEP) since PEP has a higher negative DG of phosphate hydrolysis than ATP.
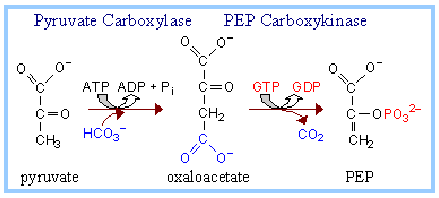
In Gluconeogenesis,
The two enzymes that catalyze the reactions for the bypass of the Pyruvate Kinase reaction are the following:
a) PEP Carboxylase Reaction
Pyruvate + HCO3– + ATP –> Oxaloacetate + ADP + Pi
Enzyme: Pyruvate Carboxylase
(b) PEP Carboxykinase Reaction:
Oxaloacetate + GTP –> Phosphoenolpyruvate + GDP + CO₂
Enzyme: PEP Carboxykinase
Contributing to the spontaneity of the two-step pathway are the following:
- The free energy of cleavage of one ~P bond of ATP is conserved in the carboxylation reaction. Spontaneous decarboxylation contributes to the spontaneity of the second reaction (PEP synthesis).
- Cleavage of a second ~P bond of GTP also contributes to driving the synthesis of PEP.
About Biotin Vitamin:
Pyruvate carboxylase utilizes biotin as a prosthetic group.
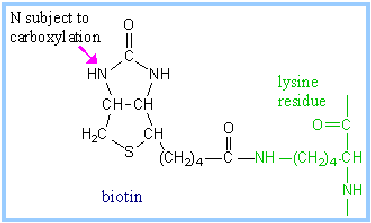
- Biotin has a 5-carbon side chain whose terminal carboxyl is in an amide linkage to the e-amino group of lysine of the enzyme.
- The biotin and lysine side chains together form a long swinging arm that allows the functional group of biotin to swing back and forth between two active sites.
- Biotin carboxylation is catalyzed at one active site by pyruvate carboxylase.
- ATP reacts with HCO₃⁻ to yield carboxy phosphate. The carboxyl is transferred from this ~P intermediate to the N of a ureido group of the biotin ring system.
Overall:
Biotin + ATP + HCO₃⁻ –> carboxy-biotin + ADP + Pi
Biotin-dependent enzymes in animals
| Enzyme | Role |
| Pyruvate Carboxylase | The first reaction in a pathway that converts 3-carbon precursors to glucose (gluconeogenesis) |
| Acetyl-CoA carboxylase | Commits acetate units to fatty acid synthesis by forming malonyl-CoA. |
| Propionyl-CoA Carboxylase | Converts propionate to succinate, which can then enter the citric acid cycle. |
| beta-Methylcrotonyl-CoA carboxylase | Catabolism of leucine and certain isoprenoid compounds |
Other Reactions in Gluconeogenesis
1. Pyruvate Carboxylase
The enzyme converts pyruvate to oxaloacetate, which is allosterically activated by acetyl coenzyme A. The adaptive value of this regulation relates to the interconnectedness of the pathways, as shown at right.
Acetyl CoA enters the Krebs cycle by condensing with oxaloacetate, whose concentration tends to be limiting for the Krebs cycle. When gluconeogenesis is active in the liver, oxaloacetate is diverted to form glucose (via PEP).
Oxaloacetate depletion hinders acetyl-CoA entry into the Krebs cycle. The resulting increase in [acetyl CoA] activates pyruvate carboxylase to synthesize more oxaloacetate.
2. Lactate to Glucose
The major breakdown product of anaerobic glycolysis in muscle is lactic acid. In muscle tissue, it’s called lactic acid. Muscle tissue is, however, not capable of re-synthesizing glycogen from lactate.
This conversion, therefore, takes place entirely in the liver. Muscle lactate is transported by the blood to the liver, where it is converted into glucose and glycogen by enzymes involved in gluconeogenesis.
Liver glycogen then breaks down into glucose and is carried back to the muscles by the blood. This conversion of muscle lactic acid to glucose in the liver and its re-entry into muscle is called the Cori cycle.
3. Amino acids to Glucose
The major portion of glucose formed in gluconeogenesis comes from amino acids. Glycogenic amino acids are converted into either citric acid cycle intermediates or pyruvate. These substances come from glucose in the liver.
Pyruvate is carboxylated to form OAA by pyruvate carboxylase and ATP in mitochondria. Further reactions in the formation of glucose take place in the cytoplasm, and therefore, oxaloacetate must come out of the mitochondria.
Oxaloacetate, however, does not readily permeate through the mitochondrial membrane and thus requires conversion to a compound that could diffuse out of the mitochondria. This is achieved mainly by its conversion to malate, which readily passes through the mitochondrial membrane. In the cytoplasm, malate is reconverted to oxaloacetate.
Oxaloacetate is then decarboxylated and phosphorylated to form phosphoenolpyruvate (PEP) by phosphoenolpyruvate carboxykinase (PEPCK) using GTP. The conversion of phosphoenolpyruvate (PEP) to fructose-1, 6-diphosphate is carried out by enzymes of glycolysis found in all tissues.
The hydrolysis of fructose diphosphate to form fructose-6-phosphate requires a specific fructose diphosphatase. Fructose-6-phosphate also requires a specific glucose-6-phosphatase for its conversion to glucose. Both the abovementioned specific enzymes are found only in liver and kidney tissues.
4. Glycerol to Glucose
Glycerol arising from the breakdown of triacylglycerides is also a good source for the synthesis of glucose in the liver. It requires initial phosphorylation by ATP followed by reduction to form DHAP (dihydroxyacetone phosphate), which enters the pathway of gluconeogenesis.
Importance of Gluconeogenesis
- A continual supply of glucose is necessary as a source of energy, especially for the nervous system and the erythrocytes.
- A gluconeogenesis mechanism is used to clear the products of the metabolism of other tissues from the blood, e.g., lactate, produced by muscle and erythrocytes, and glycerol, which is continuously produced by adipose tissue.
How is glycogenolysis regulated?
- Glucagon and glucocorticoids increase NeoGlucogenesis.
- Insulin inhibits the process.
- When glucagon levels rise, the cellular cAMP levels also increase. The cAMP concentration inactivates the enzyme pyruvate dehydrogenase (PDH) by a phosphorylation process. This would ensure that more pyruvate is converted to oxaloacetate and thereby channel neoglucogenesis.
- Ethanol inhibits this pathway.
- Gluconeogenesis and glycolysis operate in opposite directions; their regulatory signals are reciprocal to prevent futile cycling.
The summary has been given in the below table
| Enzyme | Activation | Inhibition |
| Pyruvate Carboxylase | Cortisol, Glucagon, Adrenaline, Acetyl CoA | Insulin, ADP |
| PEPCK | -do- | Insulin |
| Fru-1,6-bis-Phosphate | -do- | Fru-1,6-BisP, AMP, Fru-2,6-BP |
| Glc-6-Phosphatase | -do- | Insulin |
What is gluconeogenesis, and where does it occur?
Gluconeogenesis is the metabolic pathway by which the body synthesizes glucose from non-carbohydrate precursors such as lactate, amino acids, and glycerol. It primarily occurs in the liver (approximately 90%) and, to a lesser extent, in the renal cortex (about 10%).
During prolonged fasting or starvation, gluconeogenesis becomes the primary source of blood glucose as glycogen stores are depleted.
This pathway is critical for maintaining glucose homeostasis and supporting glucose-dependent tissues like the brain and red blood cells.
The need to use glucose is paramount, so a large body directly converts fat and ketogenic amino acids into glucose-6P using gluconeogenesis or glycolysis (which happens in the liver during prolonged fasts) thanks to the hexokinase enzyme.
Since it is called “glucose coming out,” it has nothing to do with glucose coming out of the cells—it’s about generating more glucose for fuel during starvation periods.
If a mammal without insulin cannot produce anything other than pyruvate (also known as dihydroxyacetone phosphate) from fat breakdown, its body is headed towards gluconeogenesis and ketoacidosis.
How do you know if gluconeogenesis is working?
Gluconeogenesis can be assessed through blood glucose levels and metabolic markers. During fasting, a stable blood glucose level (70–99 mg/dL) indicates that gluconeogenesis is functioning normally.
Clinically, insulin suppression tests and isotope-tracer studies (using labeled gluconeogenic precursors like ¹³C-lactate) are used to measure gluconeogenic flux directly.
In diabetic patients, elevated fasting blood glucose often signals dysregulated, overactive gluconeogenesis in the liver.
Many people struggling with excess body fat may manage this by eating less, mainly because we can’t store more than a certain amount of stored energy in our adipose tissue, which is an accumulation of the fat that accumulates from food.
It might be wise to reduce your calories instead, rather than stocking up on extra energy to overcompensate and give you weight control problems down the line.
Frequently Asked Questions (FAQs) on Gluconeogenesis
What is the gluconeogenesis pathway?
The gluconeogenesis pathway is the anabolic process that synthesizes glucose from non-carbohydrate precursors such as lactate, glycerol, and glucogenic amino acids. It mainly occurs in the liver and partly in the kidneys. The pathway reverses most steps of glycolysis but uses specific bypass enzymes to overcome the irreversible glycolytic reactions and maintain glucose homeostasis.
What is the rate-limiting step of gluconeogenesis?
The rate-limiting step of gluconeogenesis is the conversion of fructose-1,6-bisphosphate to fructose-6-phosphate, catalyzed by fructose-1,6-bisphosphatase. This enzyme is strongly regulated by energy status and hormonal signals. High ATP and citrate levels stimulate it, whereas AMP and fructose-2,6-bisphosphate inhibit it, helping coordinate gluconeogenesis with glycolysis
What is the difference between glycogenolysis and gluconeogenesis?
Glycogenolysis is the breakdown of stored glycogen to release glucose, mainly in the liver and muscle, providing a rapid source of energy. Gluconeogenesis, in contrast, generates new glucose from non-carbohydrate precursors like lactate, glycerol, and amino acids, especially during fasting when glycogen stores are depleted. Both processes help maintain blood glucose levels.
How is pyruvate converted to oxaloacetate in gluconeogenesis?
In gluconeogenesis, pyruvate is converted to oxaloacetate inside the mitochondria by the biotin-dependent enzyme pyruvate carboxylase. This reaction uses ATP and requires acetyl-CoA as an allosteric activator. Oxaloacetate then serves as a key intermediate that can be further converted toward phosphoenolpyruvate and ultimately to glucose.
How is oxaloacetate converted to phosphoenolpyruvate (PEP)?
Oxaloacetate is converted to phosphoenolpyruvate (PEP) by the enzyme phosphoenolpyruvate carboxykinase (PEPCK). This decarboxylation and phosphorylation step uses GTP as an energy source. It is a crucial bypass reaction in gluconeogenesis, allowing the pathway to proceed past the irreversible pyruvate kinase step of glycolysis.
What is the gluconeogenesis cycle in biochemistry?
The gluconeogenesis cycle describes the multi-step sequence that converts pyruvate or other precursors into glucose, often linked with the Cori cycle and alanine cycle. Lactate or alanine from peripheral tissues returns to the liver, is converted back to glucose via gluconeogenesis, and then sent again to those tissues for energy production.
How does biotin function in gluconeogenesis?
Biotin acts as a coenzyme for carboxylase reactions in gluconeogenesis, particularly pyruvate carboxylase, which converts pyruvate to oxaloacetate. It carries activated carbon dioxide during the carboxylation step. Adequate biotin is therefore essential for efficient glucose production from non-carbohydrate precursors in the liver and kidney.
What are the key enzymes of the gluconeogenic pathway?
Key enzymes in the gluconeogenic pathway include pyruvate carboxylase, phosphoenolpyruvate carboxykinase (PEPCK), fructose-1,6-bisphosphatase, and glucose-6-phosphatase. These enzymes bypass the irreversible steps of glycolysis and are major control points. Their activity is regulated by hormones such as insulin, glucagon, and cortisol, as well as cellular energy levels.
Clinical Significance
- In the enzyme pyruvate carboxylase, a deficiency is seen as an inborn error of metabolism, where mental retardation is manifested.
- Its incidence is one in every 25,000 births.
- The pyruvate carboxylase gene is located on human chromosome No. 11.
- In type II diabetes mellitus conditions, the rise in gluconeogenesis is responsible for the production of excess glucose after an overnight fast.
Final summary
Gluconeogenesis is an essential anabolic pathway that synthesizes glucose from non-carbohydrate precursors such as lactate, glycerol, and glucogenic amino acids, mainly in the liver and kidney cortex.
By bypassing the irreversible steps of glycolysis through key enzymes like pyruvate carboxylase, PEPCK, fructose‑1,6‑bisphosphatase, and glucose‑6‑phosphatase, it helps maintain normal blood glucose levels during fasting, prolonged exercise, and starvation.
Understanding its substrates, steps, regulation and differences from glycogenolysis and glycolysis is fundamental for interpreting metabolic adaptation in health and disease.
Discover more from Biochemistry Den
Subscribe to get the latest posts sent to your email.