Electron Transport Chain: Components, Mechanism, and Inhibitors in Mitochondria
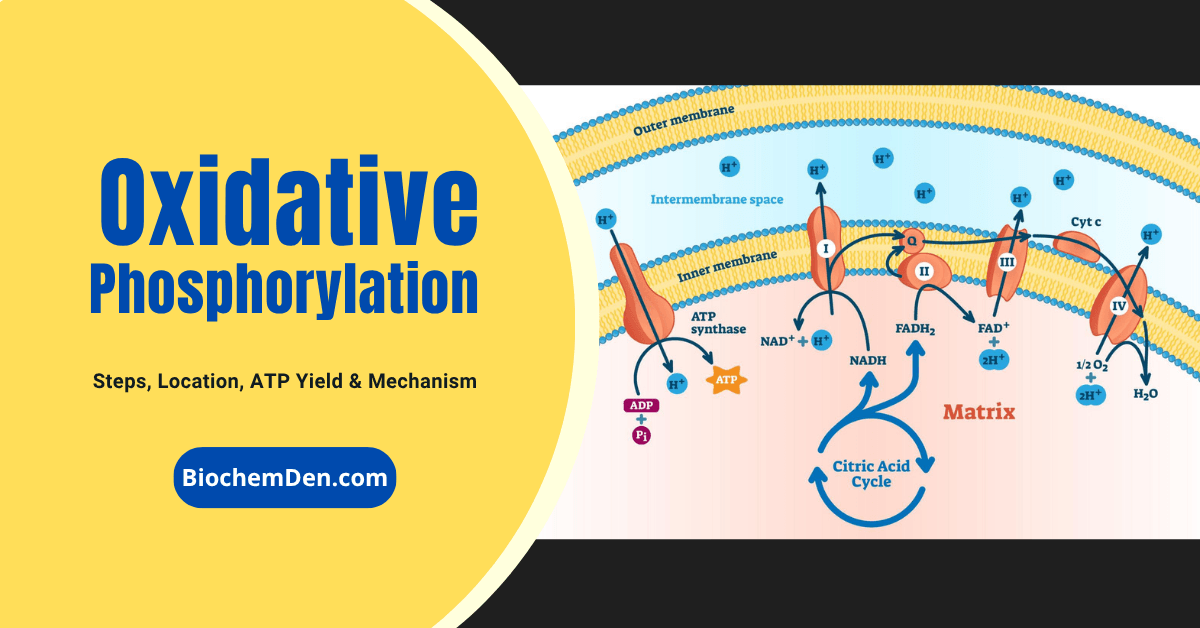
The electron transport chain (ETC) is the final and most energy-producing stage of cellular respiration, playing a critical role in ATP production in aerobic organisms. Located in the inner mitochondrial membrane, this process uses high-energy electrons from NADH and FADH₂ to drive a series of redox reactions that ultimately generate a proton gradient. This gradient powers ATP synthase, producing the majority of adenosine triphosphate (ATP), the primary energy currency of the cell.
As a key component of oxidative phosphorylation, the electron transport chain is essential for efficient energy metabolism and is tightly linked to oxygen consumption, making it fundamental to understanding how cells produce energy under aerobic conditions. Disruptions in the ETC can lead to reduced ATP yield and are associated with metabolic disorders, mitochondrial diseases, and the action of certain toxins and inhibitors such as cyanide.
In this comprehensive guide, we’ll explore the structure and function of the electron transport chain, including the roles of Complex I, II, III, and IV, the mechanism of electron flow, chemiosmosis, and ATP generation. We’ll also examine ATP yield calculations, the role of oxygen as the final electron acceptor, and clinically relevant inhibitors that affect cellular respiration.
What Is the Electron Transport Chain?
The electron transport chain is a series of membrane-embedded supramolecular protein complexes that transfer electrons from electron donors (such as NADH and FADH₂) to a final electron acceptor—molecular oxygen (O₂)—ultimately producing water.
This process is coupled to the pumping of protons across the inner mitochondrial membrane, generating an electrochemical gradient that drives ATP synthesis via the ATP synthase complex (chemiosmotic theory).
Gentle treatment of the inner mitochondrial membrane with detergents allows the separation of four distinct electron-carrier complexes (Complex I–IV), each capable of catalyzing electron transfer through a specific portion of the chain:
- Complex I and II transfer electrons to ubiquinone from NADH and succinate, respectively.
- Complex III transfers electrons from ubiquinone to cytochrome c.
- Complex IV transfers electrons from cytochrome c to molecular oxygen.
Electron Transport Chain Components
Five main types of electron carriers participate in the mitochondrial ETC, along with copper ions (Cu²⁺) that function in cytochrome oxidase.
1. Nicotinamide Nucleotides (NAD⁺ / NADH)
NAD⁺ (nicotinamide adenine dinucleotide) is the primary electron acceptor in catabolic reactions. In the TCA cycle, oxidations involving malate and isocitrate directly transfer two hydrogen equivalents to NAD⁺.
For reactions catalyzed by pyruvate dehydrogenase and α-ketoglutarate dehydrogenase, electrons pass first to lipoic acid and then via a flavoprotein to NAD⁺.
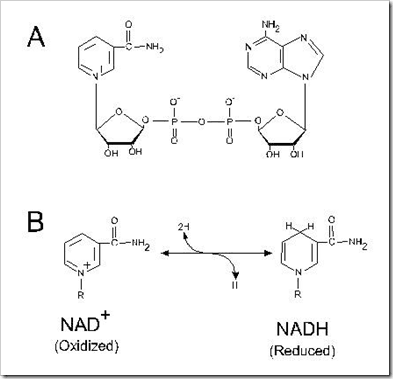
Reaction:
SH₂ + NAD⁺ → S + NADH + H⁺
NADH then donates its electrons to Complex I to begin the respiratory chain.
2. Flavoproteins (FMN and FAD)
Flavoproteins contain a tightly—sometimes covalently—bound flavin nucleotide, either FMN (flavin mononucleotide) or FAD (flavin adenine dinucleotide).
Unlike NAD⁺, the standard reduction potential of a flavin nucleotide depends on the protein with which it is associated, making the flavin nucleotide an integral part of the enzyme’s active site rather than a freely diffusible cofactor.

An important property of flavoproteins is their ability to accept one or two electrons, allowing them to serve as intermediates between reactions that donate two electrons (like NADH) and those that accept only one.
Key reactions:
NADH + H⁺ + FMN → NAD⁺ + FMNH₂
Succinate + FAD → Fumarate + FADH₂
3. Iron–Sulfur (Fe-S) Proteins
Iron–sulfur proteins were first identified as ferredoxin, a reducing agent in nitrogen fixation and photosynthesis in plants, before their role in the mitochondrial electron transport chain was recognized. Their key features include:

- Iron atoms arranged in pairs within an iron–sulfur bridge, coordinated by the sulfur atoms of cysteine residues in the protein.
- Some Fe-S proteins contain only two iron atoms (Fe₂S₂), while others contain four (Fe₄S₄)—as seen in spinach ferredoxin.
- They participate in single-electron transfer steps throughout the ETC, especially within Complexes I, II, and III.
4. Quinones (Ubiquinone / Coenzyme Q)
Mitochondria contain a specialized quinone called ubiquinone (also known as Coenzyme Q or simply “Q”), a benzoquinone with a long hydrophobic isoprenoid side chain. Its versatility makes it a critical hub in the ETC:
- It can accept one electron to form the semiquinone radical (QH•), or two electrons to form ubiquinol (QH₂).
- Because it is small and hydrophobic, it is freely diffusible within the lipid bilayer of the inner mitochondrial membrane, allowing it to shuttle reducing equivalents between less mobile electron carriers.
- Because it carries both electrons and protons, it plays a central role in coupling electron flow to proton movement across the membrane (critical for the Q cycle in Complex III).
5. Cytochromes
Cytochromes are proteins with strong absorption of visible light due to their iron-containing heme prosthetic groups. The term “cytochrome” comes from the Greek for “cellular color.” Mitochondria contain three main classes:

| Type | α-Band (reduced) | Key Properties |
|---|---|---|
| Cyt. a | ~600 nm | Found only in mitochondria; part of cytochrome oxidase |
| Cyt. b | ~563 nm | Integral membrane protein; part of Complex III |
| Cyt. c | ~550 nm | Water-soluble; mobile electron shuttle between Complex III and IV |
In all cytochromes, the iron atom in the heme group cycles between Fe³⁺ and Fe²⁺ states to transfer single electrons. The heme group is bound non-covalently in Cyt. a and Cyt. b, but in Cyt. c it is covalently attached via two thioether linkages to cysteine residues of the apoprotein.
Cytochrome a and a3 (Cytochrome Oxidase)
- Molecular weight: 72,000–93,000 Da; oxidation potential: +0.29 V.
- Both Cyt. a and Cyt. a3 contain the same heme a iron–porphyrin complex, but they are located at different positions within the apoprotein.
- Heme a is paired with one copper ion and functions as the anaerobic oxidizing unit.
- Heme a3 is paired with a second copper ion (CuB) at the O₂ binding site on subunit I, functioning as the aerobic reducing unit.
- Cyt. a absorbs at 605, 517, and 414 nm; Cyt. a₃ absorbs at 600 and 445 nm.
- Cyt. a3 (together with CuB) constitutes the terminal oxidase of the cytochrome chain.
Cytochrome b
- Contains protoporphyrin IX, but with a different apoprotein structure.
- α-absorption near 563 nm; β-absorption near 530 nm; γ-absorption near 430 nm.
- Oxidation potential: +0.04 V (in mitochondria); –0.34 V (free form).
- Thermostable and not easily extractable from the membrane.
- Does not react with O₂, CO, or CN⁻.
- Reduced by accepting an electron from ubiquinol (CoQH₂).
Cytochrome c
- The best-characterized of all cytochromes; water-soluble and easily extractable.
- α-absorption near 550 nm; β near 521 nm; γ near 416 nm.
- Contains one polypeptide chain of 104 amino acids.
- Acts as an electron carrier because its iron readily changes valence: Fe³⁺ + e⁻ → Fe²⁺.
- Unlike integral membrane complexes, Cyt. c is a freely soluble protein that moves between Complex III and Complex IV, collecting electrons from one and delivering them to the other.
- Importantly, Cyt. c also plays a role in apoptosis (programmed cell death), where its release from the mitochondria signals the activation of caspases.
Protein Complexes of the Mitochondrial ETC
The electron carriers described above do not act as isolated molecules scattered randomly across the membrane. Instead, they are precisely organized into four large, membrane-embedded supramolecular complexes (Complexes I–IV), each of which occupies a specific functional position along the electron transport chain.
These complexes were identified by gently treating the inner mitochondrial membrane with detergents, which resolved them into separable, catalytically active units—a landmark discovery that confirmed the modular architecture of the respiratory chain.
Organization Within the Inner Mitochondrial Membrane
All four complexes are integral proteins of the inner mitochondrial membrane (IMM), one of the most protein-dense biological membranes known. The IMM is highly impermeable to ions and small molecules—a property that is essential for maintaining the electrochemical gradient that powers ATP synthesis.
Its characteristic folded extensions, called cristae, dramatically increase the surface area available for housing these complexes, allowing mitochondria to maximize their ATP output.
The four complexes do not work in rigid physical contact with each other. Instead, two mobile carriers—ubiquinone (Coenzyme Q) and cytochrome c—shuttle electrons between them:
- Ubiquinone is lipid-soluble and diffuses laterally within the lipid bilayer, ferrying electrons from Complexes I and II to Complex III.
- Cytochrome c is a small, water-soluble protein that moves along the outer face of the IMM, transferring electrons from Complex III to Complex IV.
This arrangement gives the ETC both flexibility and efficiency—electrons from multiple donors can funnel through a common pool of ubiquinone before proceeding to Complex III.
Supercomplexes (Respirasomes)
In recent decades, structural studies using cryo-electron microscopy have revealed that Complexes I, III, and IV do not always operate as entirely independent units.
They can physically associate into higher-order assemblies called supercomplexes or respirasomes—most commonly in the arrangement I₁III₂IV₁. These supercomplexes are thought to:
- Enhance the efficiency of electron transfer by reducing the distance electrons must travel between complexes.
- Minimize the leakage of electrons to molecular oxygen, thereby limiting the production of damaging reactive oxygen species (ROS).
- Stabilize the individual complexes within the membrane.
Complex II generally does not participate in supercomplex formation and operates independently.
Proton Pumping and the Proton-Motive Force
Three of the four complexes—Complexes I, III, and IV—function not only as electron carriers but also as proton pumps, actively translocating H⁺ ions from the mitochondrial matrix (the N-side, negative) to the intermembrane space (the P-side, positive) with each cycle of electron transfer. Complex II does not pump protons.
The net result of this pumping is the creation of a proton-motive force (PMF)—also called the chemiosmotic gradient—across the inner mitochondrial membrane. This force has two components:
- ΔpH: A chemical gradient, with the intermembrane space more acidic (higher [H⁺]) than the matrix.
- Δψ (membrane potential): An electrical gradient, with the intermembrane space carrying a net positive charge relative to the matrix.
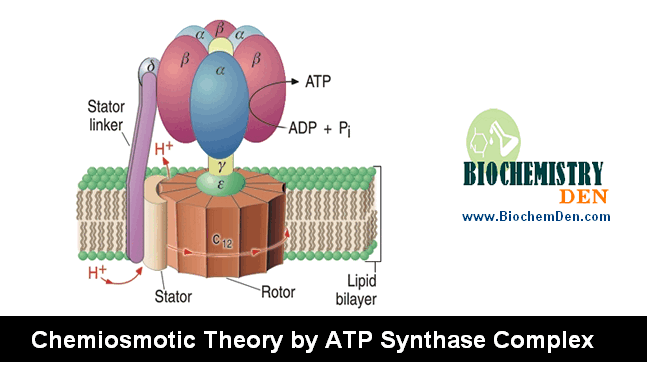
Together, these components generate a potential energy of approximately 200–220 mV across the membrane. This stored energy is harnessed by Complex V (ATP synthase), which allows protons to flow back down their electrochemical gradient through its F₀ channel, driving the rotation of its molecular rotor and coupling that mechanical energy to the synthesis of ATP from ADP and inorganic phosphate (Pᵢ).
This elegant mechanism—proposed by Peter Mitchell in his chemiosmotic theory (Nobel Prize, 1978)—explains how the free energy of electron transfer is ultimately converted into the chemical energy of ATP.
The approximate number of protons pumped per electron pair transferred is:
| Complex | Protons Pumped (H⁺/2e⁻) |
|---|---|
| Complex I | 4 |
| Complex II | 0 |
| Complex III | 4 |
| Complex IV | 4 |
| Total (NADH → O₂) | ~10 |
This stoichiometry accounts for the commonly cited yield of approximately 2.5 ATP per NADH and 1.5 ATP per FADH₂ oxidized under physiological conditions.
Electron Transport Chain Mechanism
Complex I: NADH Dehydrogenase (NADH:Ubiquinone Oxidoreductase)
Complex I — also called NADH:ubiquinone oxidoreductase — is the entry point for electrons from NADH into the respiratory chain. It is the largest complex in the ETC, composed of 42 different polypeptide chains.
Structurally, it exhibits a characteristic L-shape, with one arm embedded in the inner mitochondrial membrane and another extending into the matrix.
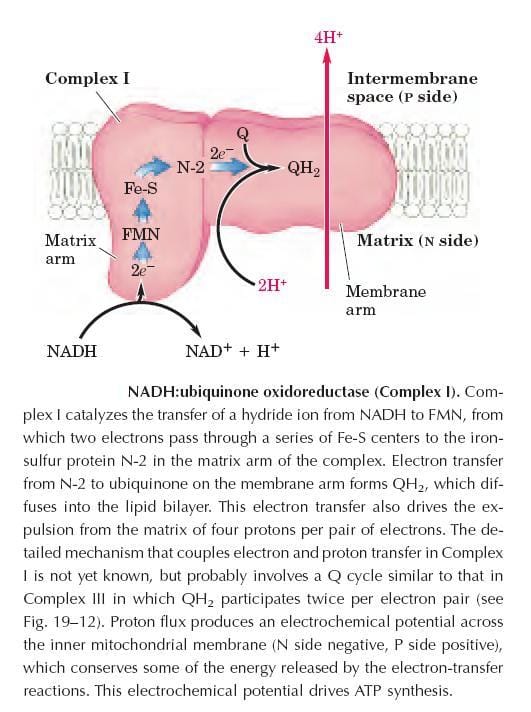
Key prosthetic groups:
- FMN (flavin mononucleotide)
- At least six iron–sulfur (Fe-S) centers
Mechanism:
- Complex I catalyzes the transfer of a hydride ion (H⁻) from NADH to FMN.
- Two electrons then pass through a series of Fe-S centers, terminating at the iron–sulfur protein N-2 in the matrix arm of the complex.
- Electrons are transferred from N-2 to ubiquinone (CoQ) located in the membrane arm, forming ubiquinol (QH₂), which diffuses freely into the lipid bilayer.
- Coupled to this electron transfer, four protons per electron pair are expelled from the matrix to the intermembrane space, contributing to the proton-motive force.
Electron flow summary:
NADH → FMN → (Fe-S₁) → (Fe-S₂) → (Fe-S₃) → (Fe-S₄) → CoQ
Energy released: –12 kcal/mol. This drives the synthesis of one ATP molecule via ATP synthase.
The exact mechanism coupling electron and proton transfer in Complex I is still being studied but likely involves a Q cycle similar to that in Complex III, in which QH₂ participates twice per electron pair.
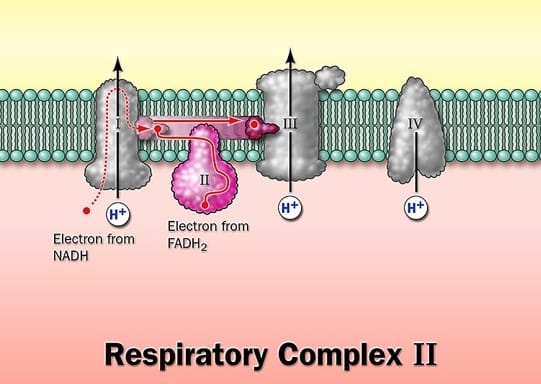
Complex II: Succinate Dehydrogenase
Complex II catalyzes the reduction of ubiquinone (CoQ) using electrons removed from succinate, directly linking the citric acid cycle to the respiratory chain. It is the only membrane-bound enzyme of the TCA cycle.
Reaction:
Succinate + CoQ → Fumarate + CoQH₂
Structure: Composed of four polypeptide subunits (molecular weights: 70,000 / 27,000 / 15,000 / 13,000 Da). Contains:
- A covalently bound FAD and an Fe-S center (4 Fe atoms) in one subunit
- A second iron–sulfur protein subunit
Mechanism: Electrons pass from succinate to FAD, then through the Fe-S centers to ubiquinone.
Note: Complex II does not pump protons across the membrane, so it contributes less energy to ATP synthesis than Complexes I, III, or IV. Other mitochondrial dehydrogenases — such as acyl-CoA dehydrogenase and glycerol-3-phosphate dehydrogenase — also feed electrons into ubiquinone at this level. Their reducing equivalents enter via the electron-transferring flavoprotein (ETF):ubiquinone oxidoreductase pathway, bypassing Complex II entirely.
Complex III: Ubiquinone–Cytochrome C Oxidoreductase (Cytochrome bc₁ Complex)
Complex III couples the transfer of electrons from ubiquinol (QH₂) to cytochrome c with the vectorial transport of protons from the matrix to the intermembrane space. It is a multiprotein complex containing
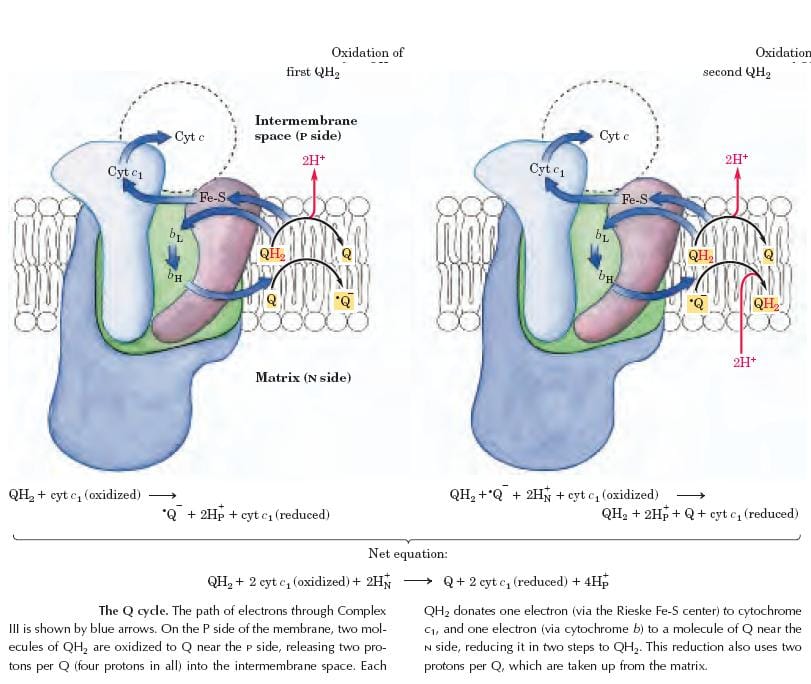
- A cluster of iron–sulfur (Fe-S) proteins (Rieske protein)
- Cytochrome b (Cyt. b)
- Cytochrome c1 (Cyt. c1)
Both Cyt. b and Cyt. c1 contain heme prosthetic groups. During electron transfer, iron in the heme group cycles between Fe³⁺ and Fe²⁺ states.
Overall reaction:
QH₂ + Cyt. c (oxidized) → Q + Cyt. c (reduced)
Mechanism — The Q Cycle:
The Q cycle is the mechanism by which Complex III achieves net proton pumping. In each cycle, two molecules of QH₂ are oxidized, and one molecule of Q is reduced. For every pair of electrons delivered to Cyt. c, four protons are translocated to the intermembrane space (two from QH₂ oxidation + two from Q reduction on the matrix side).
This amplification effect makes Complex III particularly efficient in contributing to the proton-motive force.
Energy released: –10 kcal/mol → one molecule of ATP synthesized.
Complex IV: Cytochrome Oxidase
Complex IV is the terminal complex of the respiratory chain, transferring electrons from reduced cytochrome c to molecular oxygen (O₂), reducing it to water (H₂O). It is tightly integrated into the inner mitochondrial membrane and also functions as a proton pump.
Structure:
- Contains two heme groups (heme a and heme a₃) and two copper ion centers (CuA and CuB).
- The functional unit is a single protein referred to as cytochrome a,a3.
- The three proteins most critical to electron flow are subunits I, II, and III; the complete complex consists of 13 subunits total.
Mechanism:
- Two molecules of reduced Cyt. c each donate one electron to the binuclear CuA center (subunit II).
- Electrons pass from CuA through heme a to the binuclear Fe-Cu center formed by Cyt. a3 and CuB (subunit I).
- Molecular oxygen binds to heme a3 and is reduced to its peroxy derivative (O₂²⁻) by two electrons from the Fe-Cu center.
- Delivery of two more electrons from Cyt. c converts the peroxy intermediate to two molecules of water, consuming four “substrate” protons from the matrix.
- Simultaneously, four additional protons are pumped from the matrix by a mechanism not yet fully understood.
Summary:
4 Cyt. c (reduced) + O₂ + 4 H⁺ (matrix) → 4 Cyt. c (oxidized) + 2 H₂O
Energy released: –24 kcal/mol → one molecule of ATP synthesized.
Path of Electrons Through Complex IV
The electron flow through Complex IV proceeds as follows:
Cyt. c → CuA → Heme a → Heme a3 / CuB (Fe-Cu center) → O₂ → H₂O
Four electrons are accepted sequentially from cytochrome c. The partial reduction intermediates (superoxide, peroxide) are kept tightly bound within the complex to prevent the release of reactive oxygen species. The complete four-electron reduction of O₂ to 2 H₂O requires four protons from the mitochondrial matrix as substrates, and four more protons are pumped across the membrane for every O₂ reduced.
Inhibitors of the Electron Transport Chain
The transfer of electrons through the ETC can be selectively blocked at various points by a variety of chemical agents. Understanding these inhibitors is important both for biochemistry education and for pharmacological applications, as some serve as drugs or insecticides.
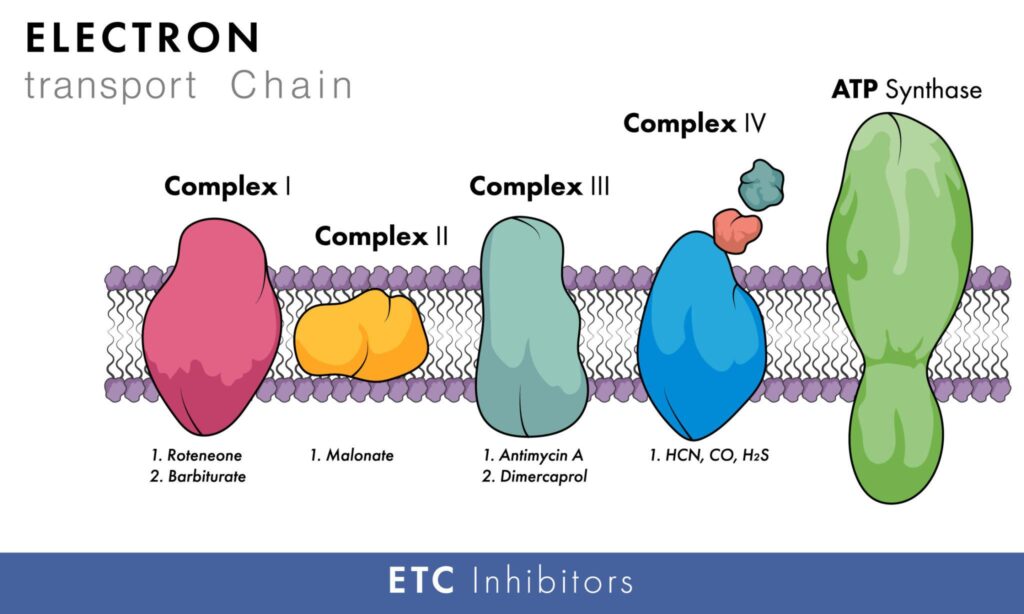
Complex I Inhibitors
| Inhibitor | Notes |
|---|---|
| Rotenone | Fish poison and insecticide; blocks electron transfer through Complex I (NADH–Q reductase) |
| Amobarbital (Amytal) / Secobarbital | Barbiturate sedatives inhibit electron transfer by competing with CoQ |
| Piericidin A | Antibiotic that blocks electron transfer by competing with CoQ |
| Chlorpromazine | Antipsychotic drug with Complex I-inhibiting properties |
| Guanethidine | Antihypertensive drug that inhibits Complex I |
Complex II Inhibitors
| Inhibitor | Notes |
|---|---|
| Carboxin | Fungicide; specific inhibitor of succinate dehydrogenase |
| TTFA (Thenoyltrifluoroacetone) | Chelator that inhibits the Fe-S cluster of Complex II |
| Malonate | Competitive inhibitor of succinate dehydrogenase (competes with succinate) |
Complex III Inhibitors
| Inhibitor | Notes |
|---|---|
| Antimycin A | Antibiotic inhibits the Q cycle by blocking electron transfer from heme b_H to ubiquinone |
| BAL (Dimercaprol) | Heavy metal antidote that also inhibits Complex III |
| Phenformin | Biguanide antidiabetic drug; hypoglycemic agent that inhibits Complex III |
Complex IV Inhibitors
| Inhibitor | Notes |
|---|---|
| Cyanide (CN⁻) | Binds tightly to Fe³⁺ in heme a3, preventing O₂ binding |
| Hydrogen Sulfide (H₂S) | Inhibits cytochrome oxidase at high concentrations |
| Azide (N₃⁻) | Similar mechanism to cyanide; binds to heme a3 |
| Carbon Monoxide (CO) | It competes with O₂ at the O₂-binding site on Cyt. a3; inhibition can be reversed by illumination with light |
Summary: The Big Picture of Electron Flow
The complete pathway of electrons through the ETC can be summarized as:
NADH → Complex I → Ubiquinone (CoQ) → Complex III → Cytochrome c → Complex IV → O₂ (→ H₂O)
FADH₂ (succinate) → Complex II → Ubiquinone (CoQ) → Complex III → …
At each major step, protons are pumped across the inner mitochondrial membrane (at Complexes I, III, and IV), building up the proton-motive force that drives ATP synthesis by ATP synthase (Complex V).
This overall process—linking electron transport to ATP production—is the basis of oxidative phosphorylation, the most efficient energy-producing pathway in aerobic life.
Discover more from Biochemistry Den
Subscribe to get the latest posts sent to your email.